
Introduction
Germination process has been widely recognized as an effective, affordable, and simple biochemical technique for enriching the nutritional profiles, bioactive compounds, and potential health benefits of the seeds. Especially, gamma-aminobutyric acid (GABA), an adequate inhibitory neurotransmitter in mammalian brains, is reported to be yielded and accumulated in the seeds during germination 1,2. Furthermore, the essential amino acids, which improve lean body mass and strength as well as are considered major materials for the generation of brain neurotransmitter 3, were also found to significantly increase in buckwheat grains during germination 4. These enhancements occur as the action of endogenous enzymes, which were formed and activated during seed germination 5,6. For instance, glutamate decarboxylase (GAD) is accountable for forming GABA from L-glutamic acid via a decarboxylation reaction. Furthermore, stressful environments during germination strongly contributed to diverse changes in the physicochemical compositions of seeds 7. Notably, acidified water for soaking is one of the key stress stimuli to induce seedling development and benefit nutritional profiles and GABA contents of germinated seeds, and it is currently researched widely. Songtip et al. 8 reported that soaking seeds in acidic water significantly impacted the GABA accumulation of germinated brown rice grains. Xu et al. 9 revealed that the citric acid-pretreated method of adlay seeds exhibited a considerable increase in free amino acids (FAA), phenolic, and flavonoid contents. Furthermore, the increment in reducing sugar, B vitamins, crude protein, and antioxidant activity of rice beans (Vigna umbellata) under acid pre-treatment were also found by Sritongtae et al. 10.
Mung bean seeds (MBs) (Vigna radiata L.) are among the most valuable pulses owing to its available abundance of essential amino acids, protein (25-28 %), fiber, vitamins, and minerals, as well as negligible lipid content (1-2 %) 11. Additionally, it is acknowledged as a meat analogue in human diets concerning its nutritional value, affordability, and digestibility 11. It is also gaining popularity as a valuable raw ingredient for formulating healthy products. However, raw mung bean seeds possess meager concentrations of GABA 12. The employment of acidified water for soaking mung bean seeds before germinating is still widely explored because pH-adjusted solutions promote crucial fluctuations in their nutritional composition and physicochemical properties. Until now, lack of study on the changes in GABA levels and nitrogen-containing constituents of MBs pretreated with pH-adjusted soaking water before germinating has been found in the literature. Therefore, this study aimed to evaluate the variations in the concentrations of GABA, FAA, nitrogenous constituents, and protease and GAD activities of MBs immersed in acidified water with diverse pH values and germinated for different times.
Materials and methods
Materials
Mung bean (Vigna radiata (L.) Wilczek), a commercial product named DX208, was supplied by the Southern Seed Co., Vietnam. These grains had a moisture content of about 11% and were undamaged without any contaminants. Chemical proximate of the grains were 26.2% protein, 1.88% lipid, 3.8% ash and 68.1% total carbohydrates. Raw mung bean seeds were placed in the refrigerator before being used for germination.
All chemicals, solvents, and standard reagents of L-Leucine, GABA, and tyrosine were analytical grade and obtained from Sigma-Aldrich Chemie GmbH (Schnelldorf, Germany).
Germination of mung bean seeds
The modified technique of Yen et al. 2 was applied to germinate mung bean seeds (MBs). MBs were submerged with a defined proportion of grains and liquid (1:4, g/mL) in various pH-adjusted soaking water (pH 4.5, 5.0, 5.5, 6.0, 6.5, and 7.0, using 2.8 M citric acid) at 40 oC for 4 h. After removing the excess liquid, the immersed seeds were subsequently germinated in a sprout apparatus (Kangaroo KG-262, China). The germination process was conducted under a darkening environment at room temperature for different times from 0 to 12 h with a 2-h interval. Finally, the germinated grains were carefully taken out of the machine, peeled from the husks, dried, and ground into powder for further analysis.
Determination of nitrogen, crude protein, and free amino acid contents in germinated mung bean seeds (MBs)
The Kjeldahl procedure, which was based on the standard AOAC technique 2001.11 13, was employed to examine the total concentration of nitrogen in MBs. This value was then utilized to estimate the crude protein content by multiplying with 6.25 as the nitrogen-to-protein conversion factor.
Protein and non-protein nitrogen concentrations were analyzed using the modified method of Wongsiri et al. 14. MBs powders (15 g) were first blended with 100 mL of distilled water, and subsequently stirred at 45 oC for 1 h before centrifuging to get rid of the residue. Afterwards, the supernatant was precipitated by 15 % aqueous trichloroacetic acid (v/v) and centrifugated to collect the solid part. The obtained residue was utilized to measure the protein nitrogen content by applying the Kjeldahl approach, while the value of non-protein nitrogen was computed by subtracting the concentration of protein nitrogen from the total nitrogen content.
The adapted procedure described by Tian et al. 15 was implemented to quantify the amount of FAA in germinated MBs. The mixture was first prepared from 5 g of MBs powder and 50 mL of 10 % acetic acid (v/v), and then incubated at ambient temperature for 1 h, before centrifugation (4,000 g, 10 min). Sodium acetate buffer (pH 5.4) was subsequently added into the achieved supernatant to 50 mL. Afterwards, the diluted sample (2 mL) was reacted with ninhydrin to form a colorimetric solution, which was analyzed at 580 nm. The calculation of FAA content was based on the calibration curve plotted with L-Leucine as a standard reagent.
Evaluation of protease activity in germinated mung bean seeds (MBs)
The separation of crude proteases was conducted following the approach of Rani et al. 16 with modifications. The mixture of powdered MBs (2 g) and chilled phosphate buffer (50 mM, pH 7.5) was first prepared, mixed well, then incubated at 4 ℃ for 30 min, and finally passed through a vacuum filtration apparatus to remove the solid part. Subsequently, the filtrate went through the centrifugation step (4,000 g, 4 ℃, 10 min) to collect the filtered liquid, which was further applied to measure protease activity.
The proteolytic assay of the germinated MBs was conducted according to the modified technique of Sattar et al. 17. The obtained extract (1 mL) containing protease was mixed well with 1 mL of 1 % casein solution (w/v, prepared in the buffer solution of pH 7.5). The content was then placed at 37 ℃ for 30 min, subsequently mixed with 2 mL of 10% aqueous trichloroacetic acid (v/v), and finally kept for 10 min to terminate the reaction. Afterwards, the supernatant collected from the centrifugation of the whole mixture was used to estimate the release of tyrosine via the Folin reaction. Particularly, the mixture was prepared from the centrifugated extract (1 mL), sodium carbonate solution (0.4 M, 5 mL), and Folin-Ciocalteu reagent (1 mL). The whole combination was placed at 37 ℃ for 20 min, followed by measuring at 660 nm by the spectrophotometer. The estimation of tyrosine released was based on the calibration curve achieved with tyrosine as a standard reagent. Furthermore, protease activity (1 U) was represented as the enzyme concentration causing the liberation of 1 μg tyrosine per minute on a mg dry weight basis.
Investigation of GABA content in the germinated mung bean seeds (MBs)
Amounts of GABA were estimated by a colorimetric technique according to the procedure of Yen et al. 2. The mixture was first prepared by MBs and ethanol (70%, v/v) in a ratio of 1:20 (g/mL), shaken continuously at 40 ℃ for 1 h, and finally centrifuged at 3,500×g to collect the supernatant. The solid part was again extracted with the same amount of ethanol (70%, v/v) twice more. Afterwards, all liquid parts from three times of separation were combined and evaporated at 45 ℃ to collect the final extract. After evaporation, a mixture was prepared from 3 mL of the diluted extract, 0.2 mL of 0.2 molL-1 borate buffer (pH 9.0), 1.0 mL of 6.0% phenol reagent, and 0.4 mL of 9.0% sodium hypochlorite. The whole mixture was mixed well, boiled for 10 min, rapidly cooled down, made up to 10 mL, and analyzed at 645 nm using the spectrophotometer. The GABA content was calculated from the standard graph and expressed as milligrams of GABA per l kg of sample.
Evaluation of glutamate decarboxylase activity in germinated mung bean seeds (MBs)
The adapted procedure of Zhang et al. 18 was applied to evaluate the GAD activity of MBs. The extraction of the GAD enzyme was conducted by mixing 5 g of germinated mung bean powder and 50 mL of chilled buffer (pH 5.5), reacting for 1 h, performing vacuum filtration, and finally centrifuging the filtrate at 4,000×g for 10 min. Subsequently, the extract solution (3 mL) was blended with 6 mL of buffer (pH 5.5) containing 1% sodium glutamate, 0.2 mmolL-1 Pyridoxal-5-Phosphate, and 1 mL of distilled water. The combination was incubated at 40 ℃ for 1 h. Then, the reaction was terminated at 90 ℃ for 5 min, centrifuged to collect the supernatant, and finally analyzed to determine the amount of GABA liberated. GAD activity (1 U) was detailed as the enzyme concentration causing the liberation of 1 μmol of GABA per hour at 40 ℃ on a gram dry weight basis.
Statistical analysis
All values attained were the average of triplicate performance. SPSS software (version 20, SPSS Inc., USA) was used to interpret and compare the results (p < 0.05) with the aid of analysis of variance and Duncan’s post-hoc multiple comparisons.
Results and Discussion
Changes in GABA content and GAD activity in MBs during germination
Figure 1 and Table 1 demonstrate the change in the level of GABA and GAD activity in MBs soaked in different acidified water and germinated for 12 h, respectively. The results showed that a gradual increment in the concentration of GABA in MBs during germination for the first 8 h was observed. The GAD activities were also found to be vigorously activated and rapidly increased during germination in this duration regardless of any pH values from 4.5 to 7.0. However, a longer germination time of over 8 h caused a slight reduction in GABA level of MBs. This could be explained by the catalyzed performance of the endogenous transaminase or other GABA-degradation enzymes to break down GABA during extended germination time 18,19. Previous studies also reported that GAD acts as a key enzyme in GABA synthesis, resulting in a positive correlation between GAD activity and GABA contents 18-20.
Regarding the influence of acidified water, both GABA content and GAD activity in MBs at any germination stage considerably improved along with lessening the pH value from pH 7.0 to pH5.5 and reached a peak at pH 5.5. Thus, a pH range of 4.5 – 6.0 was implied as a suitable pH range for high GAD activity and GABA production with the most suitable pH value of 5.5. These findings coincided with previous studies that recorded a similar range of pH values for high GAD activity in germinated faba beans 21 or paddy rice 22. The pH5.8 was also found to be the optimum value for the GABA production in foxtail millet 23. The slight differences in the optimal pH value for the action of the GAD and the accumulation of GABA might be due to different cultivars, grain storage technology, pretreating methods, and germination conditions. In addition, the seeds enhanced the formation of GABA as an adaptive reaction to stress-induced challenges 24,25. The acidic condition might be responsible for the decline in pH values of the cytosol, leading to a significant stimulation of endogenous enzymes, involving the GAD, and finally a great production of GABA 24. However, an excess reduction in pH (from pH5.0 to pH4.5) might act as a deterrent to H+ consumption during decarboxylation 24, resulting in a decrease in GAD activity as well as GABA accumulation.
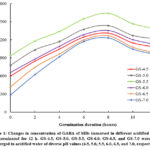 |
Figure 1: Changes in concentration of GABA of MBs immersed in different acidified water and germinated for 12 h. |
Table 1: Change in glutamate decarboxylase (GAD) activities (U/g, db) of MBs immersed in different acidified water and germinated for 12 h1
|
Germination time (hours) |
pH-adjusted value |
|||||
|
4.5 |
5.0 |
5.5 |
6.0 |
6.5 |
7.0 |
|
|
0 |
20.1cA ± 0.1 |
27.5eA ± 0.1 |
36.4fA ± 0.1 |
24.1dA ± 0.1 |
16.7bA ± 0.1 |
13.9aA ± 0.1 |
|
2 |
25.1cB ± 0.1 |
33.3eB ± 0.2 |
42.5fB ± 0.1 |
29.4dB ± 0.1 |
21.2bB ± 0.2 |
18.0aB ± 0.1 |
|
4 |
30.9cC ± 0.2 |
39.5eC ± 0.2 |
49.4fC ± 0.1 |
35.3dC ± 0.2 |
26.5bC ± 0.1 |
23.1aC ± 0.1 |
|
6 |
37.1cE ± 0.2 |
46.5eE ± 0.1 |
57.4fE ± 0.1 |
42.3dE ± 0.1 |
32.2bE ± 0.2 |
27.5aE ± 0.2 |
|
8 |
39.8cG ± 0.3 |
49.8eG ± 0.3 |
60.9fG ± 0.3 |
45.9dG ± 0.1 |
34.9bG ± 0.3 |
30.7aG ± 0.3 |
|
10 |
38.4cF ± 0.3 |
48.1eF ± 0.1 |
59.0fF ± 0.2 |
43.2dF ± 0.2 |
32.9bF ± 0.2 |
28.1aF ± 0.1 |
|
12 |
36.5cD ± 0.1 |
45.0eD ± 0.1 |
56.7fD ± 0.2 |
40.1dD ± 0.1 |
29.5bD ± 0.2 |
24.2aD ± 0.1 |
|
1Means values followed by the different capital character in the same column or the different small character in the same row are statistically different (p < 0.05). |
||||||
Changes in free amino acid (FAA) content and protease activity in MBs during germination
FAA content and protease activity in MBs steeped in the acidified water with diverse pH values and germinated for 12 h are presented in Figure 2 and Table 2, respectively. Regardless of any pH values from 4.5 to 7.0, The amount of FAA tended to boost significantly for the first 8 h, peaked at the 8-hour of germination, and then began to lessen slightly for the next 4 h, whereas protease activities were more activated with prolonging germination time. The proteolytic cleavage of proteases could explain this increment in the concentrations of FAA. However, these monomers might be exploited to synthesize new proteins or enzymes to support the new seedling, so a longer germination time of over 8 h induced a decline in these contents.
Furthermore, pH value of soaking water also affected the changes in free amino acid contents and protease activity of germinated seeds. According to Table 2, the protease activity of mung bean seeds at any germination stage remarkably intensified along with dropping pH value from pH7.0 to pH5.5, reached the highest performance in soaking water of pH5.5 and finally reduced with the further acidic solution. Furthermore, the amounts of free amino acids released were greatly proportional to the protease activity intensity due to its proteolytic cleavage ability. Hence, FAA content of MBs at any germination stage shared a similar tendency to protease activity when the steeping water was acidified from pH7.0 to pH4.5. MBs, which were first immersed in water of pH 5.5 and then germinated for 12 h, had the highest protease activity (3.11 U/mg, db) compared to other germinated ones in this study, resulting in the highest free amino acid contents (5.25 g/kg, db). Thus, this result indicated that the most favorable pH value for activating proteolytic activity in the germinated mung bean seeds was pH5.5. These results were also agreeable to the observations of Sritongtae et al. 10, who affirmed that pre-treatment with citric acid before germinating induced an increment in the quantity of free amino acids in rice bean seeds over the first germination period of 12 h. This research also revealed that acidic stress stimulated the initiation of protein-coding genes for proteolytic enzymes.
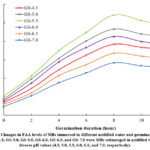 |
Figure 2: Changes in FAA levels of MBs immersed in different acidified water and germinated for 12 h. |
Table 2: Change in protease activities (U/mg, db) of MBs immersed in different acidified water and germinated for 12 h1
|
Germination time (hours) |
pH-adjusted value |
|||||
|
4.5 |
5.0 |
5.5 |
6.0 |
6.5 |
7.0 |
|
|
0 |
0.88cA ± 0.02 |
1.12eA ± 0.02 |
1.33fA ± 0.01 |
1.01dA ± 0.01 |
0.71bA ± 0.01 |
0.66aA ± 0.01 |
|
2 |
1.03cB ± 0.01 |
1.32eB ± 0.01 |
1.54fB ± 0.01 |
1.16dB ± 0.03 |
0.83bB ± 0.01 |
0.76aB ± 0.03 |
|
4 |
1.27cC ± 0.01 |
1.67eC ± 0.01 |
1.93fC ± 0.02 |
1.44dC ± 0.01 |
1.01bC ± 0.02 |
0.90aC ± 0.01 |
|
6 |
1.51cD ± 0.01 |
1.97eD ± 0.01 |
2.39fD ± 0.01 |
1.68dD ± 0.01 |
1.21bD ± 0.01 |
1.08aD ± 0.01 |
|
8 |
1.83cE ± 0.01 |
2.43eE ± 0.02 |
2.81fE ± 0.04 |
2.06dE ± 0.01 |
1.47bE ± 0.03 |
1.33aE ± 0.01 |
|
10 |
1.97cF ± 0.02 |
2.60eF ± 0.01 |
2.96fF ± 0.02 |
2.23dF ± 0.01 |
1.61bF ± 0.01 |
1.49aF ± 0.01 |
|
12 |
2.09cG ± 0.01 |
2.77eG ± 0.04 |
3.11fG ± 0.01 |
2.33dG ± 0.02 |
1.70bG ± 0.01 |
1.56aG ± 0.02 |
|
1Means values followed by the different capital character in the same column or the different small character in the same row are statistically different (p < 0.05). |
||||||
Changes in the concentrations of crude protein, protein nitrogen, and non-protein nitrogen of MBs during germination
Figures 3-5 illustrate the variations in the amounts of nitrogenous components in MBs pretreated with acidified water and germinated for 12 h. Regarding the impact of acidified water on the changes in crude protein contents of mung bean seeds at an identical germination stage, a notable reduction was witnessed along with rising pH values from pH4.5 to pH7.0. Additionally, an intensification in the pH value of the soaking water boosted from pH4.5 to pH5.5 induced an increase in non-protein nitrogen levels but a reduction in the protein nitrogen contents. When further rising pH values to pH6.0, pH6.5, or pH7.0, the former noticeably lessened compared to those at pH 5.5, while the latter had an opposite trend. The protein nitrogen contents of all mung bean seeds soaked in all acidified water markedly declined along with rising germination times while the reserved propensity was experienced with the amounts of crude protein and non-protein nitrogen. The 12-hour germinated seeds submerged in acidified water (pH 4.5) had the highest amount of crude protein (30.3%, db) and protein nitrogen (3.35%, db) among all kinds of germinated ones, while the topmost non-protein nitrogen content (1.63%, db) was obtained with soaking water of pH 5.5 and germination time of 12 h. These results were agreeable with previous studies on germinated mung bean seeds 1,26. Sritongtae et al. 10 also declared that using citric acid to acidify soaking water caused a degradation of reserve proteins in germinated rice beans. Furthermore, Xu et al. 9 revealed that the germination of adlay seeds led to the degradation of high-molecular-weight proteins due to the evolution of endogenous proteases, resulting in enhancing the quantity of low-molecular-weight proteins, peptides, and amino acids. Wongsiri et al. 14 also affirmed that the storage protein levels in the germinated mung bean seeds gradually dropped, while the non-protein nitrogen contents rose significantly under protease activation during germination time. Thus, the changes in these values were probably triggered by protease activity. During germination, protease was activated and responsible for breaking proteins down into non-protein parts like enzymes, free amino acids, peptides, or nucleic acids 27. Moreover, previous studies verified that there was a considerable increment in dry matter loss of germinated seeds 28,29, which resulted in higher protein concentrations on a unit dry basis.
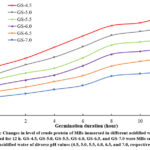 |
Figure 3: Changes in level of crude protein of MBs immersed in different acidified water and germinated for 12 h. |
 |
Figure 4: Changes in concentration of protein nitrogen of MBs immersed in different acidified water and germinated for 12 h. |
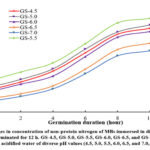 |
Figure 5: Changes in concentration of non-protein nitrogen of MBs immersed in different acidified water and germinated for 12 h. |
Conclusion
Soaking in acidified water and germination time contributed to significant changes in the concentration of GABA, FAA, and nitrogenous constituents, and protein-related enzyme activities in the mung bean seeds. For the first 8-hour germination, longer germination time induced a substantial growth in the amounts of non-protein nitrogen constituents, FAA, GABA, and protease, and GAD activities, while protein nitrogen compounds remarkably diminished. Particularly MBs immersed in acidified water of pH 5.5 and germinated for 8 h had the highest levels of GABA, free amino acids, and non-protein nitrogen. This indicated that the optimal pH of steeping water and germination time for the most effective protease and GAD activities were pH 5.5 and 8 h, respectively. Consequently, soaking raw mung bean seeds in the acidic water and then germinating them could be considered a cheap, simple, and efficient method to produce a nutrient-rich and healthy product with high medical value.
Acknowledgement
The authors sincerely thank to Vietnam National University in Ho Chi Minh City (VNU-HCM) for the research funding. We acknowledge the Ho Chi Minh City University of Technology (HCMUT), VNU-HCM for supporting our PhD student.
Conflict of Interest
The authors declare that there are no conflicts of interest related to the publication of this article.
Funding Source
This research is funded by Vietnam National University in Ho Chi Minh City (VNU-HCM) under grant number B2022-28-02.
References
- Hung PV, Yen NTH, Phi NTL, Tien NPH, Trung NTT. Nutritional composition, enzyme activities and bioactive compounds of mung bean (Vigna radiata L.) germinated under dark and light conditions. LWT. 2020;133:110100. doi:https://doi.org/10.1016/ j.lwt.2020.110100
CrossRef - Yen NTH, Hoa PN, Hung PV. Optimal soaking conditions and addition of exogenous substances improve accumulation of γ-aminobutyric acid (GABA) in germinated mung bean (Vigna radiata). https://doi.org/10.1111/ijfs.15473. Int J Food Sci Technol. 2022;57(7):3924-3933. doi:https://doi.org/10.1111/ijfs.15473
CrossRef - Rondanelli M, Opizzi A, Antoniello N, et al. Effect of essential amino acid supplementation on quality of life, amino acid profile and strength in institutionalized elderly patients. Clin Nutr (Edinburgh, Scotland). 2011;30(5):571-7. doi:10.1016/j.clnu.2011.04.005
CrossRef - Hung PV, Trinh LND, Thuy NTX, Morita N. Changes in nutritional composition, enzyme activities and bioactive compounds of germinated buckwheat (Fagopyrum esculantum M.) under unchanged air and humidity conditions. Int J Food Sci Technol. 2021;56(7):3209-3217. doi:https://doi.org/10.1111/ijfs.14883
CrossRef - Idowu AT, Olatunde OO, Adekoya AE, Idowu S. Germination: an alternative source to promote phytonutrients in edible seeds. Food Qual Saf. 2020;4(3):129-133. doi:10.1093/fqsafe/fyz043
CrossRef - Wu X, Wang Y, Tang H. Quantitative Metabonomic Analysis Reveals the Germination-Associated Dynamic and Systemic Biochemical Changes for Mung-Bean (Vigna radiata) Seeds. J Proteome Res. 2020;19(6):2457-2470. doi:10.1021/acs.jproteome.0c00181
CrossRef - Rifna EJ, Ratish Ramanan K, Mahendran R. Emerging technology applications for improving seed germination. Trends Food Sci Technol. 2019;86:95-108. doi:https://doi.org/10.1016/j.tifs.2019.02.029
CrossRef - Songtip P, Jangchud K, Jangchud A, Tungtrakul P. Physicochemical property changes in germinated brown rice flour from different storage periods of paddy rice. Int J Food Sci Technol. 2012;47:682-688.
CrossRef - Xu L, Yang N, Wu F, Jin Z, Xu X. Effect of acid pretreatment on the physicochemical and antioxidant properties of germinated adlay (Coix lachryma-jobi L.). https://doi.org/10.1111/jfpp.13511. J Food Process Preserv. 2018;42(2):e13511. doi:https://doi.org/10.1111/ jfpp.13511
CrossRef - Sritongtae B, Sangsukiam T, Morgan MRA, Duangmal K. Effect of acid pretreatment and the germination period on the composition and antioxidant activity of rice bean (Vigna umbellata). Food Chem. 2017;227:280-288. doi:https://doi.org/10.1016/j.foodchem. 2017.01.103
CrossRef - Kurek MA, Onopiuk A, Pogorzelska-Nowicka E, Szpicer A, Zalewska M, Półtorak A. Novel Protein Sources for Applications in Meat-Alternative Products – Insight and Challenges. Foods. 2022;11(7):957.
CrossRef - Ma Y, Tong L, Li J, et al. Comparison of γ-aminobutyric acid accumulation capability in different mung bean (Vigna radiata L.) varieties under heat and relative humidity treatment, and its correlation with endogenous amino acids and polyamines. Article. Int J Food Sci Technol. 2021;56(4):1562-1573. doi:10.1111/ijfs.14771
CrossRef - AOAC International. Official methods of analysis of AOAC International. Approved methods of AOAC 95046; AOAC 94205; AOAC 92039, and AOAC 200111. Gaithersburg: AOAC International Inc; 2000.
- Wongsiri S, Ohshima T, Duangmal K. Chemical Composition, Amino Acid Profile and Antioxidant Activities of Germinated Mung Beans (Vigna radiata). https://doi.org/10.1111/jfpp.12434. J Food Process Preserv. 2015;39(6):1956-1964. doi:https://doi.org/10.1111/jfpp.12434
CrossRef - Tian B, Xie B, Shi J, et al. Physicochemical changes of oat seeds during germination. Food Chem. 2010;119(3):1195-1200. doi:https://doi.org/10.1016/j.foodchem.2009.08.035
CrossRef - Rani KU, Prasada Rao UJS, Leelavathi K, Haridas Rao P. Distribution of Enzymes in Wheat Flour Mill Streams. J Cereal Sci. 2001;34(3):233-242. doi:https://doi.org/10.1006/jcrs.2000.0393
CrossRef - Sattar D-e-s, Ali TM, Hasnain A. Effect of germination on enzymatic, functional and bioactive attributes of different Pakistani legume cultivars. J Food Meas Charact. 2017;11(4):2076-2086. doi:10.1007/s11694-017-9591-5
CrossRef - Zhang Q, Liu N, Wang S, Pan L. Effects of germination and aeration treatment following segmented moisture conditioning on the γ-aminobutyric acid accumulation in germinated brown rice. Int J Agric Biol Eng. 2020;13:234-240. doi:10.25165/j.ijabe.20201305.5538
CrossRef - Luo X, Wang Y, Li Q, et al. Accumulating mechanism of γ-aminobutyric acid in soybean (Glycine max L.) during germination. I J Food Sci Technol. 2018;53(1):106-111. doi:https://doi.org/10.1111/ijfs.13563
CrossRef - Ikuta S, Fukusaki E, Shimma S. Visualization of Glutamate Decarboxylase Activity in Barley Seeds under Salinity Stress Using Mass Microscope. Metabolites.2022;12(12)doi:10.3390/metabo12121262
CrossRef - Yang R, Yin Y, Guo Q, Gu Z. Purification, properties and cDNA cloning of glutamate decarboxylase in germinated faba bean (Vicia faba L.). Food Chem. 2013;138(2):1945-1951. doi:https://doi.org/10.1016/j.foodchem.2012.11.050
CrossRef - Pramai P, Thanasukarn P, Thongsook T, Jannoey P, Chen F, Jiamyangyuen S. Glutamate Decarboxylase (GAD) Extracted from Germinated Rice: Enzymatic Properties and Its Application in Soymilk. J Nutr Sci Vitaminol. 2019;65(Supplement):S166-s170. doi:10.3177/jnsv.65.S166
CrossRef - Bai Q, Fan G, Gu Z, Cao X, Gu F. Effects of culture conditions on γ-aminobutyric acid accumulation during germination of foxtail millet (Setaria italica L.). Eur Food Res Technol. 2008;228(2):169-175. doi:10.1007/s00217-008-0920-0
CrossRef - Chung H-J, Jang S-H, Cho HY, Lim S-T. Effects of steeping and anaerobic treatment on GABA (γ-aminobutyric acid) content in germinated waxy hull-less barley. LWT. 2009;42(10):1712-1716. doi:https://doi.org/10.1016/j.lwt.2009.04.007
CrossRef - Shelp BJ, Bozzo GG, Trobacher CP, Chiu G, Bajwa VS. Strategies and tools for studying the metabolism and function of γ-aminobutyrate in plants. I. Pathway structure. Botany. 2012;90(8):651-668. doi:10.1139/b2012-030
CrossRef - Elobuike CS, Idowu MA, Adeola AA, Bakare HA. Nutritional and functional attributes of mungbean (Vigna radiata [L] Wilczek) flour as affected by sprouting time. https://doi.org/10.1002/leg3.100. Legume Sci. 2021;3(4):e100. doi:https://doi.org/10.1002/leg3.100
CrossRef - Ikram A, Saeed F, Afzaal M, et al. Nutritional and end-use perspectives of sprouted grains: A comprehensive review. Food Sci Nutr. Aug 2021;9(8):4617-4628. doi:10.1002/fsn3.2408
CrossRef - Mbithi-Mwikya S, Van Camp J, Yiru Y, Huyghebaert A. Nutrient and Antinutrient Changes in Finger Millet (Eleusine coracan) During Sprouting. LWT. 2000;33(1):9-14. doi:https://doi.org/10.1006/fstl.1999.0605
CrossRef - McCue P, Zheng Z, Pinkham JL, Shetty K. A model for enhanced pea seedling vigour following low pH and salicylic acid treatments. Process Biochem. 2000;35(6):603-613. doi:https://doi.org/10.1016/S0032-9592(99)00111-9
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








