Plasma Lipid Profiles of Transgenic Mice Expressing the Human Apob100xcetp are Altered Differentially by Diets Enriched With Defined Fatty Acids
1Faculty of Pharmacy Arab International University Damascus, Syria
2Department of Surgery University of Michigan Ann Arbor, Michigan
3College of Nursing and Allied Health Sciences Department of Nutritional Sciences Howard University Washington, DC 20059.
DOI : http://dx.doi.org/10.12944/CRNFSJ.3.3.04
Download this article as:
![]()
Dietary fat is known to modulate plasma lipid profiles. Synthesis of high density lipoproteins (HDL), which has protective effects on vascular disease is also influenced by dietary fats, but the mechanisms are unclear. The hapoB100XCETP transgenic mouse was used to investigate the effects of fatty acids on the metabolism of plasma lipoproteins, including the pathway leading to synthesis HDL. Male transgenic mice were fed with diets formulated to provide TG (33% energy) as tripalmitin (TP), triolein (TO), tristearin (TS) or equicaloric substitution of fat with carbohydrate (sucrose) for 4 weeks. Analysis of plasma profile showed that HDL-cholesterol were 53.7+14; 64.6+8.6; 50.2+3.3; 47.0+9.2 and 45.2+4.9 mg/dL for control, oleate, palmitate, stearate and sucrose based diets, respectively. LDL-cholesterol levels were 51.7+7.0; 23.1+7.0; 38.9+2.2; 75.1+1.8 and 46.8.1.0 mg/dl, for control, TO, TP, TS and sucrose, respectively. Hepatic Lecithin-cholesterol acyltransferase (LCAT) protein levels increased by 2-fold in mice fed TS or TO diets, compared to TP, while sucrose had no effect. The scavenger receptor class B type I (SR-B1) which plays an important role in meditating the uptake of HDL-derived cholesterol and cholesteryl ester in the liver and steroidogenic tissues increased in livers of animals fed TP and TO, while TS and sucrose did not have a similar effects. These results suggests that fatty acids can uniquely impact HDL, in addition, the ApoB100XCETP mouse is a useful model for the evaluation of how dietary components affect the risk of developing atherosclerosis and heart disease.
KEYWORDS:ApoB100; CETP; TC; HDL-C; LDL-C and TG; Transgenic
Introduction
The main cause of cardiovascular disease (CVD) is atherosclerosis, characterized by the combination of chronic inflammation and/or hyperlipidemia. CVD is believed to be the leading cause of death in the Western world and its prevalence is increasing in Eastern Europe and developing countries.1 High density lipoprotein-cholesterol (HDL-C) protects against vascular disease by accepting free cholesterol from macrophage foam cells in the artery wall in a process, often described as reverse cholesterol transport (RCT). Reverse cholesterol transport describes the efflux of excess cellular cholesterol from peripheral tissue and its return to the liver for excretion in the bile and ultimately the feces during digestion of dietary fat. It is believed to be a critical mechanism by which HDL exerts a protective effect on the development of atherosclerosis or vascular diseases.1,2 In the proposed paradigm, cholesterol efflux from arterial macrophages to extracellular HDL-based acceptors occurs through the action of transporters such as ATP binding cassette transporter A1 (ABCA1) and ATP-binding cassette transporter G1 (ABCG1). After efflux to HDL, cholesterol can be esterified in the plasma by the enzyme lecithin: cholesterol acyltransferase (LCAT), and is ultimately transported from HDL to the liver, either directly via the scavenger receptor BI (SR-BI) or after transfer to apoB-containing lipoproteins by the cholesteryl ester transfer protein (CETP). The physiological activator of LCAT is apolipoprotein A-I (apoA-I), the major HDL protein. However, cholesterol removal is compromised if the activity of apoA-I is compromised such as exposure to reactive intermediates.3-5
Abnormalities in HDL metabolism due to elevation in plasma TG have been reported, leading to suggestions that TG may inhibit cholesterol esterification and the binding of Lecithin-cholesterol acyltransferase (LCAT) to HDL.6 Fatty acids or TG can interfere in any of the stated pathways, and can directly or indirectly mediate activities, which result in the modulation of HDL. Our previous work suggested that oleic acid (OA) rich diets exerted a positive effect on the metabolism of plasma HDL in comparison to other fatty acids. We have previously demonstrated that dietary components such as fatty acids play key roles in the regulation of this pathway. In addition, studies demonstrated that monounsaturated fatty acids increased the expression of LCAT and SR-B1.7
Murine transgenic models expressing human genes involved inlipoprotein metabolism have increasingly served as small mammalian models,where the spectrum of both normal and pathologic human serum lipidprofiles can be simulated, and in several instances demonstrated the formation of atherosclerotic lesions.8 In the present study, the effect of fatty acids on the expression of plasma lipoproteins in general, and HDL in particular was evaluated in a transgenic animal model with a humanized lipid profile.
Materials and Methods
Materials
Semi-synthetic fats [triolein (TO), tripalmitin (TP), tristearin (TS)]used to modify the diets were a generous donation by ABITECCorporation (Columbus, OH). LCATcDNA was generously provided by Dr. R. Taramelli of Milan, Italy.The SR-B1 cDNA was a gift from Dr. Alan Tall. The antisense probe template for glyceraldehyde-3 phosphatedehydrogenase (GAPDH) was obtained from Ambion (Austin, TX)and was used to correct for RNA loading in addition to normalizingLCAT mRNA abundance. The procedure for labeling probes and measurementof LCAT mRNA abundance were previously reported (7, 9, and 10). All other chemicals were obtained from Sigma Chemical Company(St. Louis, MO).
Mouse Model
The double transgenic male mice expressing both human CETP and apoB100were obtained from Taconic Labs (Germantown, NY) and bred to obtain sufficient animals to conduct this study. Tail biopsy for DNA analysis and genotyping were carried out when the mice were 13 days old, followed by immersion of the tail in ice cold ethanol for 10 seconds to minimize pain. These mice arehemizygous for both transgenes and are on a mixed (C57BL/6xSJL) background. In this animal, the LDL-cholesterol is increased by three to four fold and the HDL-cholesterol is reduced approximately by 60% as compared to non-transgenic control mice. The percentages of the total cholesterol within the HDL, LDL, and VLDL fractions of the hApoB100XCETP animals are approximately 30%, 65%, and 5% respectively, similar to the distribution of cholesterol in the plasma of normolipidemic human profile [non-HDL to HDL of 2:1. The non-transgenic mice have a non-HDL to HDL ratio of 1:4.11
At 6 weeks of age, the mice (18 to 20 g. wt.) were placed on diet in which greater than 90% of the total fatty acids were a single fatty acid species. The control diet was a commercially available mouse chow to which the synthetic fats were added to provide 30% total calories from fat. The synthetic fats consisted of triolein (TO, oleic acid), tripalmitin (TP, palmitic acid), and tristearin (TS, stearic acid). One diet was formulated to provide the same amount of calories from sucrose (Su). Concomitant with the diet shift from normal laboratory chow to the defined fatty acid one, mice were randomly divided into 5 experimental groups according to previous treatments and fed the respective diets for 7 weeks. All the protocols for animal use were approved by Wayne State University Institutional Animal Investigation Committees and the study was conducted in accordance with the Guide for the Care and Use of Laboratory Animals (U.S. Department of Health and Human Services: National Research Council, 1985).
Serum Lipoprotein Analysis
Lipoprotein cholesterol distribution was determined by agarose gel electrophoresis ([Helena laboratory, Beaumont, TX].12, 13 This method was compared to the Fast protein liquid chromatography (FPLC) technique and the results were found to be greater than 95% correlated. Total cholesterolwas quantified enzymatically (Sigma) from theindividual sera as well as from 100-ul aliquots of the FPLCfractions. The relative amount of cholesterol within each peakwas determined by area quantitation under the curve using the appropriate baseline modifications from the FPLC cholesteroltracings. Serum triglyceride levels were measured by enzymaticassay (Sigma). Lipoprotein cholesterol distribution was determined by agarose electrophoresis (Helena Laboratory, Beaumont, TX).14,15
Immuno-analyses of hepatic scavenger receptor class B, type 1 receptor protein.
Liver SR-BI receptor protein mass determination was carried out as reported previously.7 Twenty five µg of solubilizedliver membrane protein was subjected to protein electrophoresisusing 4–12% pre-cast polyacrylamide gradient gels undernon-reducing conditions (Novex, San Diego, CA). Proteins were transferred onto nitrocellulose membranes and incubated overnightwith a 1:2000 dilution of mouse SR-B1 antibody (Novus Biologicals,Littleton, CO) and visualized by chemiluminescence detection(Alpha Innotech, San Leandro, CA). Quantification of the specificluminescent protein bands was performed with Image-Quant software(Molecular Dynamics, Sunnyvale, CA).
Immuno-analyses of hepatic Lecithin: cholesterol acyltransferase.
Mouse liver was prepared by homogenizing 0.2g of liver in 5mL of [150 mmol/L NaCl, 1 mmol/L CaCl2, 10 mmol/LTris- HCl (pH 7.5); and the protease inhibitors, pH 7.5] (Lyses buffer). The supernatant was prepared by centrifugingthe homogenate at 8000 x g for15 min. Supernatant protein concentrationswere determined by the Bio-Rad dye-binding protein assay.
For LCAT protein immunoblotting, 25 µg of solubilizedliver protein was subjected to protein electrophoresisusing 12% polyacrylamide gels undernonreducing conditions. Proteins weretransferred onto nitrocellulose membranes and incubated overnightwith a 1:2000 dilution of rLCAT anti rabbit antibody (developed in-house) and visualized by chemiluminescence detection(Alpha Innotech, San Leandro, CA). Quantification of the specificluminescent protein bands was performed with Image-Quant software(Molecular Dynamics, Sunnyvale, CA).
Measurement of LCAT/SRr-B1 mRNA.
Poly (A+) RNA was isolated from the liver according to the Trizolreagent protocol (Invitrogen kit; Invitrogen, Carlsbad, CA).Northern blot analysis was performed to quantitate mRNA levelsof LCAT. For each sample, 5 µg of RNA was electrophoresedthrough formaldehyde- 1.2% agarose gels and transferred to 20xSSC-equilibrated Hybond ECL nitrocellulose membranes. Membraneswere hybridized after UV crosslinking and washed at a high stringency(65°C, 0.1x SSC). Northern blots were probed with labeled([32P]dCTP, specific activity > 3000 Ci/mmol) as previously described (9). The levels of LCAT/SR-B1 mRNA in liversamples were was normalized to glyceraldehyde phosphate dehydrogenase(GADPH) mRNA levels for each sample.
Results
Characterization of the hApoB100/CETP mouse
The lipid profile of the hApoB100/CETP mouse was characterized by measuring its lipid profile (Figure 1) and compared it to the wild type, when fed a regular chow diet. As shown in Figure 3, the LDL-C of these mice is increased three to four fold and the HDL-cholesterol reduced by approximately 60% as compared to non-transgenic control mice. The percentages of the total cholesterol within the HDL, LDL, and VLDL fractions of the ApoB100xCETP animals were confirmed and found to be similar to the distribution of cholesterol in the plasma of normolipidemic humans.11 Thus, by expressing both human apoB and human CETP, the lipoprotein cholesterol distribution in the serum of a chow-fed mouse was transformed into one that resembles a human profile (non-HDL to HDL of 2:1).
 |
Figure 1: FPLC cholesterol elution profile of transgenic for the human apoB100xCETP mice and control (wildtype C57BL/6) mice, indicating the relative positions of VLDL LDL and HDL.
|
Cholesterol profile as determined by agarose electrophoresis and confirmed by FPLC.
The profiles shown (Figure 2) are representative of the distribution of cholesterol in the various lipoproteins from each dietary group. Total cholesterol did not differ between the groups. However, animals fed TO had significant elevation in HDL-C when compared to control. Tristearin fed animals on the other hand had higher, while triolein had lower, LDL-C when compared to control.
 |
Figure 2: Representative lipoprotein-cholesterol profile of male hApoB100xCETP transgenic mice fed different fatty
|
Body weights and serum lipids of mice fed the control diet and diets with different triglycerides
Final body weights of mice and relative liver weights did not statistically differ by diet or among groups. As shown in Table 1, serum LDL-C levels were significantly (p<0.05) lowerin mice fed the triolein diet, while LDL-C levels were higher in mice fed tristearin compared to control group. As was observed in our previous rat study,7,9 animals that were fed triolein had higher HDL-C content and reduced LDL-C levels. Although the distribution of cholesterol in HDL for the other treatments appeared similar, there was a tendency to have reduced HDL-C levels with sucrose and tristearin compared to tripalmitin. Reduced HDL-C with tripalmitin is not what is often seen in human. However, the decrease in HDL-C was also accompanied by reduced LCAT expression suggesting that tripalmitin had a negative impact on the esterification of cholesterol into HDL, in part by down-regulation of LCAT. The feeding of sucrose generated the highest TG concentration and the lowest HDL-C levels as expected, though not statistically different (Table 1). Additionally, there were differences seen in the level VLDL-C in agreement with numerous studies that FA also contribute to plasma cholesterol (other than HDL), and contributes to the atherogenic process as well. The ratio of TC: HDL-C was higher than control in all the groups except for triolein, while LDL: HDL-C and TG: HDL-C ration were higher than control only in mice fed the tristearin and sucrose diets (P<0.05) (Table 2). Even though Apo A-I was higher in the triolein group when compared to the other diet groups, no difference was observed between the treatments for Apo A-II. In addition, hepatic lipase activity levels were not different among the groups (Table 2).
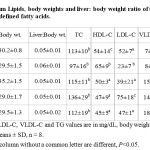 |
Table 1: Serum Lipids, body weights and liver: body weight ratio of transgenic mice fed fats with defined fatty acids. Click here to View table |
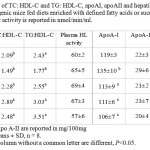 |
Table 2: Ratio of TC: HDL-C and TG: HDL-C, apoAI, apoAII and hepatic lipase activity levels of transgenic mice fed diets enriched with defined fatty acids or sucrose Plasma hepatic activity is reported in nmol/min/ml. Click here to View table |
Expression of LCAT protein in hepatic tissue
The results showed that triolein increased LCAT protein levels. In addition, mice fed tristearin diets, had hepatic LCAT protein levels that were significantly higher compared to mice fed control diet (Figure 3). The elevated protein levels in the previous groups contrasted with a significant down regulation in the tripalmitin fed group. To determine if the increase in LCAT protein was a function of induction of the LCAT gene, we looked at the levels of messenger RNA in the liver. The mRNA levels correlated with the protein levels in the triolein group, however LCAT mRNA levels did not correlate with its protein levels for the tripalmitin treated animals.
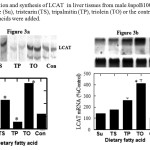 |
Figure 3: Induction and synthesis of LCAT in liver tissues from male hapoB100xCETP transgenic mice fed sucrose (Su), tristearin (TS), tripalmitin (TP), triolein (TO) or the control diet (Con) to which the fatty acids were added.
|
Hepatic SR-B1 receptor
The hepatic scavenger receptor B1 protein concentration wasincreased nearly 2.5-fold in mice fed the triolein and tripalmitin diets compared tomice fed with control diet (P < 0.05) (Figure 4). The observed increase in SR-B1 protein levels following exposure to triolein and tripalmitin was accompanied by a similar fold induction of SR-B1 mRNA (Figure 5). No similar induction was observed with other tristearin or sucrose.
The level of LCAT in 25 ug of liver protein extracts was determined by western blot analysis using an antibody raised against a synthetic peptide to LCAT (A). The relative level of LCAT protein in the liver were quantified by UV absorbance and normalized based on protein loaded (D). Induction of hepatic LCAT mRNA by fatty acids is shown in (B). The level of LCAT mRNA in liver samples was normalized based on GAPDH levels as internal control for each sample. The data shown in the graph (C) represent normalized values of LCAT mRNA abundance expressed as a percent of Control fed mice. Each point represent mean +/- SE of the mean for data obtained from 4 animals/group
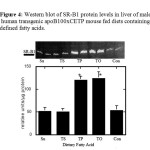 |
Figure 4: Western blot of SR-B1 protein levels in liver of male human transgenic apoB100xCETP mouse fed diets containing defined fatty acids. |
Protein extracts of the liver were obtained from animals fed a mouse chow diet enriched with sucrose (Su), tristearin (TS), tripalmitin (TP) triolein (TO) or the chow control diet (Cc). 25ug of nuclear extracts were determined by western blot analysis using ECL detection kit and mouse antibody. SR-B1 protein levels in the livers are expressed as relative to total hepatic protein loaded. Each point represent mean +/- SE for data from 5 animals/group. *(p<0.05)
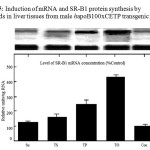 |
Figure 5: Induction of mRNA and SR-B1 protein synthesis by fatty acids in liver tissues from male hapoB100xCETP transgenic mice. |
The level of SR-B1 mRNA in liver samples was normalized based on GAPDH levels as internal control for each sample. The data shown in the graph (C) represent normalized values of SR-B1 mRNA abundance expressed as a percent of Control fed mice. Each point represent mean +/- SE of the mean for data obtained from 4 animals/group.
Discussion
The main objective of this study was to obtain a better understanding of how DIETARY fatty acids influence HDL-C concentration using an animal model, genetically altered to incorporate human genes in the RCT pathway (human ApoB100xCETP transgenic mice with a lipid profile similar to humans). The use of this transgenic model permits a clinical evaluation of the changes at the hepatic LCAT and SR-B1 mRNA and protein levels and subsequent effects on blood lipids (TC, HDL-C and LDL-C). Low plasma concentrations of HDL-C and high VLDL/ LDLC are independent risk factors for the development of atherosclerosis.1 In addition, the ratio of VLDL/LDL to HDL is to a great extent affected by the cholesteryl CETP.50 When these animals are placed on an atherosclerotic diet, they display a significant decrease in HDL compared to wild animals. Using this approach, the effects of feeding diets enriched in different fatty acids can be evaluated. Initially, the animals were characterized by measuring its lipid profile and comparing it to the wild type, when fed a regular chow diet. As shown in Figure 1, the LDL-cholesterol of these mice is increased three to four fold and the HDL-cholesterol decreased approximately 60% as compared to non-transgenic control mice. The distribution of total cholesterol within the HDL, LDL, and VLDL fractions of the hapoB100xCETP animals were similar to the distribution of cholesterol in the plasma of normolipidemic humans.
Contrary to the rat model,11 feeding these fatty acids in the genetically altered mouse did not affect total cholesterol. However, the different fatty acid treatments modulated HDL, LDL and VLDL-C differently. The triolein fed group, for example, had a significant increase in HDL-C that was accompanied by a significant decrease in LDL-C. The current observation that plasma HDL-C levels tend to increase when oleic acid is fed in the diet confirms our previous report,7 which might explain in part, observations in population studies illustrating the benefits of the Mediterranean diet that is known to be rich in olive oil.13,14 Mice fed the tristearin diet had significant increase in plasma LDL, VLDL-C and TG levels. This observation contradicts assertions that dietary stearic acid is neutral14,15 in its cholesterol raising effects. Similarly, our finding is not in agreement with the notion that palmitic acid increased total cholesterol and LDL cholesterol.16 As expected, the sucrose fed group had increased TG and VLDL-C concentration, with no changes in HDL or LDL cholesterol concentrations compared to control.
The increase in HDL-C levels observed in the triolein group was accompanied by an increase in Apo A-I concentration. Other dietary treatments failed to show any increase in Apo A-I levels. Epidemiological studies revealed an inverse relationship between coronary artery disease and the plasma Apo A-1 concentration.17,18,19 Thus, our results fits the general scheme of what has been described as the protective effect of diets rich in oleic acid. However, caution has to be exercised when interpreting these data, since the increase in Apo A-1 might be accompanied by an increase in Apo A-II, leading to a decrease in Apo A-I/Apo A-II ratio. In fact, several studies in mice and in humans demonstrated an association between increased levels of apoA-II levels and increased susceptibility to atherosclerosis,20-21 increased free fatty acid levels.20,22 obesity,and increased metabolic syndrome.23,24 Hedrick et al.,25 suggested that as the ratio of apoA-II to apoA-I increases, HDL particles become larger because of the inhibition of hepatic lipase (HL), leading to the loose of the anti-atherogenic properties of HDL. Taking this into consideration we measured Apo A-II and Apo A-I/Apo A-II ration, and the results showed no statistical difference among the triolein group and the other including control group; thus, eliminating apoA-II as a confounding factor when evaluating the effects of fatty acid on RCT in particular and on the lipid profile in general.
The role of hepatic lipase (HL) in the metabolism of HDL is well established.26 HL acts on decreasing HDLcore lipids, apparently making lipid-poor apoA-I available fordissociation or transfer to other particles.26-28 In addition, HL is thought to enhance hepatic HDL cholesterol uptake.29,30 Thus, an increase in HDL-C level may be due to a decrease in hepatic lipase activity and vice versa. This notion is supported by Homanics et al.,31 who reportedthat HL-deficient mice have a mild dyslipidemia with increased cholesterol, phospholipid, and large HDLfloating in the 1.02-1.04 g/ml density range. The results from this study showed no difference in hepatic lipase activity between the dietary treatments. This is contrary to reports by Privett and colleagues32 using rats that were essential fatty acid deficient. In addition, their dietary treatment included of trans-fatty acids, which are more likely to have different metabolic pathways than fatty acids in the cis isoform.
Hepatic Scavenger receptor B1 is an important factor in modulating serum HDL levels. In vivo and in vitro studies have supported its participation in the selective uptake of HDL cholesterol33-37 and involved in RCT. A 2-fold increase in SR-B1 protein was observed following exposure to triolein accompanied by a similar fold induction of SR-B1 mRNA. The amount of SR-B1 in liver protein extracts from TP fed mice where almost just as elevated as those on triolein, with corresponding increases in its mRNA. It is interesting to note that although induction of SR-B1 was observed in the mice fed both triolein and tripalmitin, no effect was observed in the tristearin fed or the sucrose fed group. Spady et al.,38 reported that saturated fats tend to decrease SR-B1 expression, however tristearin and tripalmitin are both grouped as saturated fats, but both had varying effects on SR-B1 expression. In addition, the results showed that the length of the fatty acid is not the driving force behind its effect on SR-B1. The induction of SR-B1 in the triolein group make it harder to explain the high levels of HDL-C observed in this group, because higher levels of SR-BI corresponds to lower levels of circulating HDL-C (33-37). Studies using the SR-B1–null mice confirm that changes in SR-B1 activity affect the metabolism of cholesterol and when SR-B1 is overexpressed in hepatic tissue, there is a significant decrease in plasma HDL.39 These observations do not account for plasma HDL-C levels under conditions when the reverse cholesterol transport pathway (LCAT/CETP) that gives rise to plasma HDL-C is simultaneously upregulated with the SR-B1 receptor pathways. However, it can be speculated that because SR-B1 plays an important role in meditating the uptake of HDL-derived cholesterol and cholesteryl ester in the liver and steroidogenic tissues40 the induction of SR-BI, a principal enzyme in the catabolic arm of reverse cholesterol transport, may be accompanied by a similar induction in the anabolic arm of the same pathway.
Lecithin-cholesterol acyltransferase playsa major role in RCT by esterifying free cholesterol(FC) in circulating lipoproteins to maintain a free cholesterol gradient betweenthe peripheral cells and the HDL particle surface and, accordingly,to promote FC efflux from the cells.41 LCAT from rodents and primates has different phospholipid substrate fatty acyl specificities,42,43 and they lack CETP responsible for the transfer of CE to other lipoproteins in humans. In the current study, hApoB100xCETP mice fed the TO diet had elevated levels of hepatic LCAT mRNA, compared to mice fed the tripalmitin, tristearin, sucrose or the control diets. This suggests that oleic acid may impact the expression of LCAT at some level of transcription. The levels of hepatic LCAT protein were similarly elevated in the triolein group, however a surprising increase was observed in hepatic LCAT mass in mice fed the tristearin diet. Despite this increase, HDL-C did not change. In fact, LDL-C, VLDL-C and TG were significantly higher in this group when compared to control. It is possible that the increase in cholesterol esterification by LCAT is offset by the increase in TG and VLDL-C. Similar to our previous observations7 feeding tripalmitin was accompanied by a reduction in hepatic LCAT mRNA abundance and protein mass. In fact, LCAT protein in liver extracts from TP fed mice was almost 2-fold less than that of other FAs, including the control. It is not known if this effect occurs at the level of transcription. The levels of mRNA are thought to be poor predictors of protein expression, because mRNA levels can vary almost 20 times and still yield the same level of gene products.44,45,46
Sucrose is an efficient triacylglycerol (VLDL)-elevatingagent both in humans and in animal models.47 We used it as an endogenous control for raising triglyceride, thus comparisons were made between the effects of exogenous vs. endogenous fat on RCT. Feeding sucrose did not affect total cholesterol significantly; neither did it affect hepatic LCAT; however, TG and VLDL-C were significantly higher than control. TC: HDL-C ratio was higher in mice fed sucrose, suggesting that sucrose may have a lowering effect on HDL-C. Serum HDL originates from synthesisand secretion by the liver and via the catabolism of circulatingVLDL.48,49 Decreased levels of HDL in mice fed sucrosemay be related to the decreased catabolism of VLDL, the precursorof plasma HDL, and not to the decreased LCAT production by the liver. Our data are unique in that sucrose increased serum TG but without any effect on LCAT, while it has been reported by other investigators that feeding sucrose up-regulate LCAT activity in mice and in monkeys.50,51 This might be explained by differences in the experimental models used.
A review by Hausenloy and Yellon53 suggested that low levels of high-density lipoprotein cholesterol (HDL-C; <1.03 mmol/l or <40 mg/dl) are an independent risk factor for CVD, and raising levels of HDL-C is a major treatment strategy for regressing atherosclerosis and enhancing CVD risk reduction. Accordingly, this can be achieved by both pharmacological and non-pharmacological lifestyle strategies that include dietary changes. In general, these studies agree with epidemiological data in humans suggesting that diets rich in monounsaturated fatty acids (MUFAs) do tend to lower the risk of coronary heart disease.12,13 Thus, increasing the monounsaturated fatty acyl composition of plasma CE may be beneficial in preventing atherosclerosis development.
Conclusion
In conclusion, our observations which showed that palmitate may up-regulate SR-B1 (be it in a smaller magnitude) in opposite direction to LCAT is not only surprising, but is also new and may point to a novel pathway whereby palmitic acid may contribute to the reverse cholesterol transport. It may also explain why palmitate may not elevate LDL-C as expected of a saturated FA. Our lipid data suggest that FA may play a much more significant role in the modulation of HDL-C. However, different FA species can either improve or worsen the ratio of TC: HDL-C and TG:HDL-C, resulting in a profile that impacts the risk of atherosclerosis. The above findings make it possible to speculate that the level of TG induced by these fatty acids may have a role in the uptake or turnover rates of HDL and demonstrates the continuous call for FAs to be investigated individually rather than as groups. Finally, these results suggests that not only do fatty acids uniquely impact plasma lipid profile as shown in this model, the ApoB100XCETP mouse can be a useful model for the evaluation of how diet and dietary components affect the risk of developing atherosclerosis and heart disease.
Acknowledgements
This study was a part of Dr. Hatahet’s dissertation at Wayne State University, Detroit, Michigan USA. The work was supported in part, by an institutional research fellowship and by a National Institutes of Health grant #HL03389 from the U.S. Public Health Service to Dr. Fungwe.
References
- Hansson GK. Mechanisms of disease inflammation, atherosclerosis, and coronary artery disease. N Engl J Med. 2005; 352: 1685–1695.
CrossRef - Tall AR. Plasma high density lipoproteins: metabolism and relationship to atherogenesis. J. Clin. Invest. 1990; 86: 279-284.
CrossRef - Gatto LM, Lyons MA, Brown AJ, Samman S. Trans fatty acids and cholesterol metabolism: mechanistic studies in rats and rabbits fed semipurified diets. Int. J. Food Sci. Nutr. 2001; 52: 435-341
CrossRef - Rader DJ, Alexander ET, Weibel GL, Billheimer J, Rothblat GH. The role of reverse cholesterol transport in animals and humans and relationship to atherosclerosis J Lipid Res. 2009 50, S189-S194.
CrossRef - Krieger M. Charting the fate of the “good cholesterol”: identification and characterization of the high-density lipoprotein receptor SR-B1. Annu. Rev. Biochem. 1999; 68: 523-558.
CrossRef - Skeggs JW, Morton RE. LDL and HDL enriched in triglyceride promote abnormal cholesterol transport. J Lipid Res. 2002; 43(8):1264-74.
- Hatahet W, Cole L, Kudchodkar B, Fungwe T. Dietary fats with defined Fatty acids differentially modulate serum LCAT Activity in rats: Effect on total cholesterol and triacylglycerol: High Density Lipoprotein cholesterol Ratio J. Nutrition. 2003; 133,: 689-694
- Smith JD, Breslow JL. The emergence of mouse models of atherosclerosis and their relevance to clinical research. J Intern Med. 1997; 242(2):99-109.
CrossRef - Fungwe TV, Kudchodkar BJ, Lacko AG, Dory L. Fatty acids modulate lecithin: cholesterol acyltransferase secretion independently of effects on triglyceride secretion in primary rat hepatocytes. J. Nutr. 1998; 128: 1270-1275.
- Kudchodkar BJ, Lacko AG, Dory L, Fungwe TV. Dietary fat modulates serum paraoxonase 1 activity in rats. J. Nutr. 2000; 130: 2427-33.
- Grass DS, Saini U, Felkner RH, Wallace RE, Lago WJP, Young SG, Swanson ME. Transgenic Mice expressing both human apolipoprotein B and human CETP have a lipoprotein cholesterol distribution similar to that of normolipidemic humans. J Lipid Res. 1995; 36(5): 1082-1090.
- Kris-Etherton PM, Pearson TA, Wan Y, Hargrove RL, Moriarty K, Fishell V, Etherton TD. High-monounsaturated fatty acid diets lower both plasma cholesterol and triacylglycerol concentrations. Am J Clin Nutr. 1999; 70(6):1009-15
- Kouris-Blazos A. Morbidity mortality paradox of 1st generation Greek Australians. Asia Pac J Clin Nutr. 2002; 11 Suppl 3:S569-75
CrossRef - Bonanome A, Grundy SM. Effect of dietary stearic acid on plasma cholesterol and lipoprotein levels. N Engl J Med. 1988; 318(19):1244-8.
CrossRef - Tholstrup T, Marckmann P, Jespersen J, Vessby B, Jart A, Sandstrom B. Effect on blood lipids, coagulation, and fibrinolysis of a fat high in myristic acid and a fat high in palmitic acid. Am J Clin Nutr. 1994; 60(6):919-25.
- Zock PL, de Vries JH, Katan MB. Impact of myristic acid versus palmitic acid on serum lipid and lipoprotein levels in healthy women and men. Arterioscler Thromb. 1994; 14(4):567-75.
CrossRef - Stampfer MJ, Colditz GA, Willett WC. Postmenopausal estrogen therapy and cardiovascular disease: ten-year follow-up from the Nurses’ Health Study. N Engl J Med. 1991; 325(11):756-62
CrossRef - Buring JE, O’Connor GT, Goldhaber SZ, Rosner B, Herbert PN, Blum CB, Breslow JL, Hennekens CH. Decreased HDL2 and HDL3 cholesterol, Apo A-I and Apo A-II, and increased risk of myocardial infarction. Decreased HDL2 and HDL3 cholesterol, Apo A-I and Apo A-II, and increased risk of myocardial infarction. Circulation. 1992; 85(1):22-9.
CrossRef - Amouyel P, Isorez D, Bard JM, Goldman M, Lebel P, Zylberberg G, Fruchart JC. Parental history of early myocardial infarction is associated with decreased levels of lipoparticle AI in adolescents. Arterioscler Thromb. 1993; 13(11):1640-4.
CrossRef - Warden CH, Daluiski A, Bu X, Purcell-Huynh DA, De Meester C, Shieh BH, Puppione DL, Gray RM, Reaven GM, Chen YD, et al. Evidence for linkage of the apolipoprotein A-II locus to plasma apolipoprotein A-II and free fatty acid levels in mice and humans. Proc Natl Acad Sci U S A. 1993; 90(22):10886-90
CrossRef - Mehrabian M, Qiao JH, Hyman R, Ruddle D, Laughton C, Lusis AJ. Influence of the apoA-II gene locus on HDL levels and fatty streak development in mice. Arterioscler Thromb. 1993 Jan; 13(1):1-10.
CrossRef - Escola-Gil JC, Marzal-Casacuberta A, Julve-Gil J, Ishida BY, Ordonez-Llanos J, Chan L, Gonzalez-Sastre F, Blanco-Vaca F. Human apolipoprotein A-II is a pro-atherogenic molecule when it is expressed in transgenic mice at a level similar to that in humans: evidence of a potentially relevant species-specific interaction with diet. J Lipid Res. 1998; 39(2):457-62
- Castellani LW, Goto AM, Lusis AJ. Studies with apolipoprotein A-II transgenic mice indicate a role for HDLs in adiposity and insulin resistance. Diabetes. 2001; 50(3):643-51.
CrossRef - Weng W, Breslow JL. Dramatically decreased high density lipoprotein cholesterol, increased remnant clearance, and insulin hypersensitivity in apolipoprotein A-II knockout mice suggest a complex role for apolipoprotein A-II in atherosclerosis susceptibility. Proc Natl Acad Sci U S A. 1996; 93(25):14788-94.
CrossRef - Hedrick CC, Hassan K, Hough GP, Yoo JH, Simzar S, Quinto CR, Kim SM, Dooley A, Langi S, Hama SY, Navab M, Witztum JL, Fogelman AM. Short-term feeding of atherogenic diet to mice results in reduction of HDL and paraoxonase that may be mediated by an immune mechanism. Arterioscler Thromb Vasc Biol. 2000; 20(8):1946-52.
CrossRef - Clay MA, Newnham HH, Forte TM, Barter PI. Cholesteryl ester transfer protein and hepatic lipase activity promote shedding of apo A-I from HDL and subsequent formation of discoidal HDL. Biochim Biophys Acta. 1992; 1124(1):52-8.
CrossRef - Demant T, Carlson LA, Holmquist L, Karpe F, Nilsson-Ehle P, Packard CJ, Shepherd J. Lipoprotein metabolism in hepatic lipase deficiency: studies on the turnover of apolipoprotein B and on the effect of hepatic lipase on high density lipoprotein. J Lipid Res. 1988; 29(12):1603-11
- Bamberger M, Lund-Katz S, Phillips MC, Rothblat GH. Mechanism of the hepatic lipase induced accumulation of high-density lipoprotein cholesterol by cells in culture. Biochemistry. 1985; 24(14):3693-701.
CrossRef - Taskinen MR, Packard CJ, Shepherd J. Effect of insulin therapy on metabolic fate of apolipoprotein B-containing lipoproteins in NIDDM. Diabetes. 1990; 39(9):1017-27.
CrossRef - Jansen S, Lopez-Miranda J, Salas J, Castro P, Paniagua JA, Lopez-Segura F, Ordovas JM, Jimenez-Pereperez JA, Blanco A, Perez-Jimenez F. Plasma lipid response to hypolipidemic diets in young healthy non-obese men varies with body mass index. J Nutr. 1998; 128(7):1144-9.
- Homanics GE, de Silva HV, Osada J, Zhang SH, Wong H, Borensztajn J, Maeda N. (1995) Mild dyslipidemia in mice following targeted inactivation of the hepatic lipase gene. J Biol Chem. 1995; 270(7):2974-80.
CrossRef - Privett OS, Phillips F, Shimasaki H, Nozawa T, Nickell EC. Studies of effects of trans fatty acids in the diet on lipid metabolism in essential fatty acid deficient rats. Am J Clin Nutr. 1977; 30(7):1009-17.
- Acton S, Rigotti A, Landschulz KT, Xu S, Hobbs HH, Krieger M. Identification of scavenger receptor SR-BI as a high density lipoprotein receptor. Science. 1996; 271: 518–520
CrossRef - Kozarsky KF, Donahee MH, Glick JM, Krieger M, Rader J. Gene transfer and hepatic overexpression of the HDL receptor SR-B1 reduces atherosclerosis in the cholesterol-fed LDL receptor-deficient mouse. Arterioscler. Thromb. Vasc. Biol. 2000; 20: 721-727
CrossRef - Babitt J, Trigatti B, Rigotti A, Smart EJ, Anderson RG, Xu S, Krieger M. Murine SR-BI, a high density lipoprotein receptor that mediates selective lipid uptake, is N-glycosylated and fatty acylated and colocalizes with plasma membrane caveolae. J Biol Chem. 1997; 272(20):13242-9.
CrossRef - Xu S, Laccotripe M, Huang X, Rigotti A, Zannis VI, Krieger M. Apolipoproteins of HDL can directly mediate binding to the scavenger receptor SR-BI, an HDL receptor that mediates selective lipid uptake. J Lipid Res. 1997; 38(7):1289-98.
- Temel RE, Trigatti B, DeMattos RB, Azhar S, Krieger M, Williams DL. Scavenger receptor class B, type I (SR-BI) is the major route for the delivery of high density lipoprotein cholesterol to the steroidogenic pathway in cultured mouse adrenocortical cells. Proc Natl Acad Sci U S A. 1997; 94(25):13600-5.
CrossRef - Spady DK, Kearney DM, HobbsHH. Polyunsaturated fatty acids up-regulate hepatic scavenger receptor (SR-B1) expression and HDL-cholesterol ester uptake in the Hamster. J. Lipid Res. 1999; 40: 1384-1394.
- Rigotti, A. et al . 1997. A targeted mutation in the murine gene encoding the high density lipoprotein (HDL) receptor scavenger receptor class B type I reveals its key role in HDL metabolism. Proc. Natl. Acad. Sci. USA 94: 12610–12615.
CrossRef - Valacchi G1, Sticozzi C, Lim Y, Pecorelli A. Scavenger receptor class B type I: a multifunctional receptor. Ann N Y Acad Sci. 2011. 1229:E1-7.
CrossRef - Glomset, J. A. The lecithin: cholesterol acyltransferase reaction. J Lipid Res. 1968; 9:155–167.
- Pownall HJ, Pao Q, Massey JB. Isolation and specificity of rat lecithin: cholesterol acyltransferase: comparison with the human enzyme using reassembled high-density lipoproteins containing ether analogs of phosphatidylcholine. Biochim Biophys Acta. 1985; 833(3):456-62.
CrossRef - Liu M, Bagdade JD, Subbaiah PV. Specificity of lecithin:cholesterol acyltransferase and atherogenic risk: comparative studies on the plasma composition and in vitro synthesis of cholesteryl esters in 14 vertebrate species. J Lipid Res. 1995; 36(8):1813-24.
- Chawla A, Barak Y, Nagy L, Liao D, Tontonoz P, Evans RM. PPAR-gamma dependent and independent effects on macrophage-gene expression in lipid metabolism and inflammation. Nat Med. 2001; 7(1):48-52.
CrossRef - Aebersold R, Rist B, Gygi SP. Quantitative proteome analysis: methods and applications. Ann N Y Acad Sci. 2000; 919:33-47.
CrossRef - Gygi SP, Rist B, Gerber SA, Turecek F, Gelb MH, Aebersold R. Quantitative analysis of complex protein mixtures using isotoe-coded affinity tags. Nat Biotechnol. 1999; 17(10):994-9.
CrossRef - Vrana A, Fabry P, Kazdova L, Zvolankova K. Effect of the type and proportion of dietary carbohydrate on serum glucose levels and liver and muscle glycogen synthesis in the rat. Nutr Metab. 1978; 22(5):313-20
CrossRef - Fielding C, Fielding, PE. Molecular physiology of reverse cholesterol transport. J. Lipid Res. 1995; 26: 211-228.
- Eisenberg S. High density lipoprotein metabolism. J. Lipid Res. 1984; 25: 1017-1058.
- Mattock MB, Sheorain VS, Subrahmanyam D. Lecithin-cholesterol acyltransferase activity in carbohydrate-induced hypertriglyceridemia in mice. Experientia. 1978; 34(3):304-5.
CrossRef - Sharma C, Srinivasan SR, Radhakrishnamurthy B, Berenson GS. Effect of dietary carbohydrate type on lipoprotein lipase, hepatic lipase, and lecithin:cholesterol acyltransferase activities in cynomolgus monkeys. Biochem Med Metab Biol. 1986; 36(1):51-9.
CrossRef - Ha YC, Calvert GD, Mcintosh GH, Barter PJ. A physiologic role for the esterified cholesterol transfer protein—in vivo studies in rabbits and pigs. Metabolism-Clin Exp. 1981; 30: 380–383.
CrossRef - Hausenloy DJ, Yellon DM. Enhancing cardiovascular disease risk reduction: raising high-density lipoprotein levels. Curr Opin Cardiol. 2009 Sep;24(5):473-82
CrossRef





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








