Comparative Evaluation of Plant and Whey Protein Hydrolysates: Bioavailability, Intestinal Cellular Response and Cholesterol-Regulating Gene Expression in A Caco-2 Model
 , Minakshi Dutta1, Nitin Kumar Singhal2, Rama Prashat Govindarajalu3, Navita Bansal1, Suneha Goswami1, Ranjeet Ranjan Kumar1, Aruna Tyagi1 and Vinutha Thimmegowda1*
, Minakshi Dutta1, Nitin Kumar Singhal2, Rama Prashat Govindarajalu3, Navita Bansal1, Suneha Goswami1, Ranjeet Ranjan Kumar1, Aruna Tyagi1 and Vinutha Thimmegowda1* 1Division of Biochemistry, ICAR-Indian Agricultural Research Institute, New Delhi, India
2Department of Food and Nutrition, National Agri-Food Biotechnology Institute, Mohali, India
3Division of Genetics and Plant Breeding, ICAR-Indian Agricultural Research Institute, New Delhi, India
Corresponding Author Email: vinuthabiochem@gmail.com
Download this article as:
ABSTRACT:The present study investigates the bioavailability and gene regulatory effects of hydrolyzed plant protein blends compared to hydrolyzed whey protein in Caco-2 intestinal cells, aiming to evaluate their potential in managing hypercholesterolemia. Two plant protein isolate blends derived from chickpea, sesame, peanut, and mung bean in different ratios were enzymatically digested and assessed for protein content, solubility, cytotoxicity, amino acid transport, and gene expression. Hydrolyzed whey protein isolate (WPI) showed the higher total protein and essential amino acid (EAA) bioavailability, while hydrolyzed PlPIs demonstrated higher soluble protein content and superior non-essential amino acid (NEAA) absorption. All protein hydrolysates promoted Caco-2 cell proliferation without cytotoxic effects. Gene expression analysis revealed that both WPI and plant blends hydrolysates upregulated peptide transporter PepT1 and downregulated cholesterol metabolism gene SREBF2, indicating cholesterol-lowering potential via modulation of intestinal transport and lipid metabolic pathways. While WPI showed superior EAA delivery, PlPI-blends exhibited unique NEAA profiles, with PlPI-blend 2 particularly rich in glutamic acid, arginine, and aspartic acid, the amino acids linked to cardiovascular health and metabolic regulation. These findings highlight the promise of plant protein blends, particularly PlPI-blend 2, as functional food ingredients for promoting intestinal health and regulating cholesterol metabolism.
KEYWORDS:Bioavailability; Caco-2 cell model; Cholesterol homeostasis; PepT1/SCL15A1; Plant protein isolates; SREBF2
Introduction
Hypercholesterolemia a prevalent genetic disorder marked by increased levels of low-density lipoprotein levels, significantly increasing the risk of premature cardiovascular disease, affecting approximately 1 in 200 individuals.1Effective cholesterol-lowering strategies are essential, and dietary interventions, particularly those involving functional proteins, have emerged as promising alternatives to pharmacological treatments. Proteins are essential dietary components, playing a broader role in human health beyond structural support. Consuming sufficient high-quality protein supports growth, repair, metabolism while managing chronic health conditions, positively affecting blood glucose, blood pressure and cardiovascular health, including cholesterol levels.2For example, whey protein isolate contains peptides with hypocholesterolemic and hypolipidemic effects, potentially by inhibiting cholesterol synthesis or enhancing bile acid excretion.3Plant proteins, known for their cholesterol-lowering properties, may exert beneficial effects by enhancing LDL receptor activity, promoting bile acid excretion, and modulating key genes involved in lipid metabolism.4Despite these advancements, significant research gaps remain in understanding the comparative efficacy of plant and animal-derived proteins. While animal-derived proteins like whey are well-studied for their high essential amino acid (EAA) content and bioavailability, plant proteins often lack certain EAAs but offer bioactive compounds and dietary fiber that contribute to metabolic health.5-7 Recent efforts to blend plant protein isolates (PlPIs) aim to overcome individual amino acid deficiencies and offer more complete nutritional profiles.8 However, limited research exists on how these blends influence nutrient absorption and gene expression related to cholesterol metabolism and amino acid transport. This knowledge gap hinders the optimization of plant-based proteins as sustainable dietary interventions for hypercholesterolemia. This study investigates following research questions: (1) How do two plant protein isolate blends (PlPI-blend 1 and PlPI-blend 2) compare to whey protein isolate in terms of cytotoxicity and amino acid bioavailability in the Caco-2 cell model? (2) To what extent do these protein sources modulate the expression of SLC15A1 (PepT1), a peptide transporter, and SREBF2, a regulator of cholesterol biosynthesis? The findings aim to assess the nutritional and functional potential of plant protein blends as sustainable dietary tools for managing cholesterol and supporting gut health.
Materials and Methods
Materials
Four plant protein sources (mungbean, chickpea, sesame, and peanut) and a commercial animal protein isolate (whey)were chosen for their good amino acid composition and protein content and procured from the local market in New Delhi, India. The seeds were ground into 0.5 µm flour and defatted with hexane for further analysis.
Extraction of Protein Isolates
Briefly, defatted flour was mixed with distilled water in a 1:8 (w/v) proportion and subjected to sonication. The suspension was adjusted to pH 10.0 using 2 M NaOH and stirred for 1 h at 25°C. Subsequently, it was centrifuged at 5000 × g for 15 min at 4°C, and the resulting supernatant was collected. The pH was then reduced to 4.5 with 1 M HCl to induce protein precipitation, followed by overnight incubation and a second centrifugation step under identical conditions. The precipitated proteins were rinsed twice with distilled water and neutralized to pH 7.0 using NaOH (1 M), air-dried, and ground into PlPIs.9Linear programming, a mathematical method was employed to identify the optimal composition of protein blends, with a focus on high lysine and methionine content in accordance with WHO recommendations using Microsoft Excel Solver tool. Using the results of linear programming, peanut, chickpea, sesame, and mungbean respectively were mixed in specific ratios to yield PlPI-blend 1 (0.1:0.1:0.7:0.1), and PlPI-blend 2 (0:0.1:0.8:0.1). PlPI-blend 2 didn’t contain peanut.
Protein Solubility
Total protein soluble protein of the protein isolates and blends wasquantified using the Bradford assay (Bradford,1976).10 The % protein solubility was computed as:
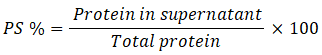
In Vitro Digestion of Protein Isolates
In brief, 200 mg sample in 2 mL water was mixedfollowed by the addition of 35 mL 0.1N HCl and overnight incubation at 37°C. Pepsin (1 mL, 3 mg/mL) was mixed with the sample, and kept for incubation(2 hours; 37°C). The pH of the digest was then neutralized to 7.4 using 1 M Tris-HCl, after which trypsin and chymotrypsin (1 mL, 2.5 mg/mL each) were added and the sample incubated for another 4 h at 37°C. Enzymatic activity was stopped by heating the mixture in boiling water for 10 minutes.11

Caco-2 Cell Culture
NCCS provided the Caco-2 cells (Pune, India) and cultured in a 37°C humidified incubator with 5% CO₂. DMEM/MEM medium enriched with fetal bovine serum (10%), L-glutamine (1%), and 1% penicillin–streptomycin was used to grow the cells. Additionally, 25 mM HEPES buffer (pH 7.4) was included to maintain pH stability. Cells were supplied with new medium every two days to maintain optimal growth. At ~80% confluence, cells were harvested using trypsin-EDTA (2.5 g/L trypsin, 0.2 g/L EDTA) and subsequently used for downstream assays.12
Cytotoxicity Assay in Caco-2 Cell
Caco-2 cells were seeded into 96-well plates and cultured under controlled conditions (37°C, 5% CO₂ atmosphere) until they achieved a density of 6 × 103 cells per well and 80% confluence. The cell layers were then rinsed with serum-free DMEM and treated with 100 μL of DMEM enriched with 10% fetal bovine serum, incorporating test samples at concentrations of 50, 100, 150, 200, and 250 μg/mL in duplicate pairs for a 24-hour incubation period.Following incubation, the plates underwent centrifugation at 1000 × g for 30 seconds to remove the old medium, followed by a PBS wash. A staining solution of 100 μL (9:1 ratio of media to MTT) was applied in darkness for 2 hours. After centrifugation, the MTT solution (0.5 mg/mL) was removed, and 100 μL of a lysis buffer (containing 8 mM HCl and 10.5% NP-40 in DMSO) was added to each well. After 5 minutes of gentle agitation, absorbance was measured at 570 nm using a Synergy H1 microplate reader (BioTek, Bad Friedrichshall, Germany).13
Bioavailability Studies and UPLC
In 6-well plates a 12,000 molecular weight cut-off dialysis membrane was fitted to the bottom of a suitably sized Transwell insert ring (polycarbonate inserts; Corning) to mimic intestinal barrier conditions. The complete setup was sterilized overnight in 0.1 N HCl at 4°C and finally, the inserts were kept immersed in sterile water for 1 hour before use.Over a 21-day culture period on Transwell inserts, Caco-2 cells developed into differentiated monolayers (~80% confluence, ~6 × 10³ cells/well). At the start of the experiment, the cell monolayers were rinsed with PBS,100 μL in vitro digested test samples (Whey, PlPI-Blend 1, and PlPI-Blend 2) were applied to the apical surface of the system (6.0 μg/μL in transport buffer, pH7).Following two hours of incubation at 37°C with 5% CO2media from both apical and basal chambers were collected.UPLC was employed to quantified total amino acids (free + peptide-derived) that was transferred to the basal side and that remained on the apical side.13
RNA–Isolation and cDNA Synthesis
Total RNA from Caco-2 cells was isolated using TRIzol reagent (Invitrogen, Carlsbad, USA). Using spectrophotometry, the isolated RNA’s content and purity were ascertained. At 260/280 nm, an optical density (OD) ratio of roughly 1.9 was regarded as suggestive of low contamination(NanoDrop, Witec, Switzerland). According to the cDNA synthesis kit’s user handbook (Thermo Scientific, USA), cDNA was created.
Quantitative real-time PCR (qRT-PCR) investigation of the expression patterns of the genes SREBF 2 (cholesterol metabolism) and SLC15A1 (amino acid transporter)
The housekeeping gene Actin1 (GenBank Accession No. NM_001101.5) served as a reference to standardize the variable expression levels of SREBF2 and SLC15A1 in Caco-2 cells. Primers were validated through conventional end-point PCR, with a melting curve analysis conducted to ensure specificity.14 For the qRT-PCR setup, 1 μL of cDNA template, 3.2 μL of sterile distilled water, 0.4 μL of each forward and reverse primer, and 5 μL of 1X SYBR Green Master Mix (Applied Biosystems, USA) were combined. A preliminary denaturation at 98°C for 10 minutes, 35 cycles of denaturation at 94°C for 50 seconds, annealing at the corresponding Ta°C for 40 seconds, and extension at 72°C for 50 seconds, and a final extension step at 72°C for 10 minutes were used to perform the real-time PCR (see Table 1 for primer details). Using the 2−ΔΔCt technique, gene expression levels were measured and reported as foldchanges in comparison to the control group.15
![]()
Table 1: Sequences of The Primers Used for Real Time PCR Amplification and The Product Size.
|
Gene name |
Primer Oligo Id | Sequence 5’ to 3’ | Amplicon length (bp) | Ta |
Accession no. |
|
SREBP2 |
HsSREBF2 FP | TTCCTGTGCCТСТССТТТААС | 142 | 62.1 | NM_004599.1 |
| HsSREBF2 RP | TCATCCAGTCAAACCAGCC |
63.8 |
|||
|
PepT1 |
HsSLC15A1 FP | CCCTGATTGTGTTTGTCCTTG | 130 | 64.2 | NM_005073.4 |
| HsSLC15A1 RP | AATGCCTTACTCCGATGCC |
63.9 |
|||
|
β-actin |
HsACTB FP | GTCTTCCCCTCCATCGTG | 120 | 63.3 | NM_001101.5 |
| HsACTB RP | GTACTTCAGGGTGAGGATGC |
61.2 |
Statistical Analysis
Each experiment was conducted in triplicate, and data are reported as mean ± SD. Group comparisons were analyzed using one-way ANOVA in SPSS version 19 (IBM, USA). Statistical significance was assessed by Student’s t-test, and differences with p < 0.05 were regarded as significant.
Results
Protein Isolates’ Solubility and Protein Content
The protein content and solubility of undigested whey protein isolate (WPI) and two formulated plant protein blends (PlPI-blend 1 and PlPI-blend 2) were evaluated (Table 2).
Table 2: Total Protein Content and Soluble Protein Content of Protein Isolates.
|
Protein isolate |
Protein content % | Soluble proteins % | In-vitro digestibility % | Total polyphenols (µg/g) |
| Whey | 82.05±0.3 | 39.5±0.9 | 86.3±0.2 |
75.03±4.3 |
|
PlPI- Blend 1 |
75.97±1.3 | 41.9±0.6 | 82.1±2.9 | 120.4±1.2 |
| PlPI- Blend 2 | 76.47±1.4 | 43.1±0.6 | 78.8±4 |
135.3±3.7 |
WPI exhibited the highest total protein content (82% w/w), whereas both PlPI-blends showed comparable values (~76% w/w). Notably, PlPI-blend 2 demonstrated the maximum soluble protein fraction (43%), followed by PlPI-blend 1 (41.8%) and WPI (39.4%). These findings suggest that although whey has higher overall protein content, plant protein blends, particularly PlPI-blend 2, offer superior solubility, a key factor influencing protein digestibility and functional application in food systems.
Effect of Plant Protein Blends On Cell Proliferation
Whey protein hydrolysate and plant protein blends(PlPI-blend 1 and PlPI-blend 2) hydrolysates significantly enhanced Caco-2 cell proliferation, as assessed by the MTT assay, which measures cellular metabolic activity. Across a range of concentrations from 50 to 250 μg/mL, none of the treatments displayed cytotoxic effects, confirming cell safety. Hydrolyzed plant protein blends demonstrated a dose-dependent boost in cell proliferation, with PIPI-blend 2 achieving the highest improvement (up to 22.8%), followed by PIPI-blend 1 (22.6%) and whey (20.6%) compared to the FBS control (Fig.1). These findings indicate thatenzymatic digestion of both plant and whey proteins yields bioactive peptides that promote intestinal cell growth, with PlPI-blend 2 showing the highest effect (22.8%), highlighting hydrolysed plant protein potential as functional food components for supporting gut health.
Fig. 1: Effect of Protein Hydrolysates (50–250 µg/ml) On The Viability of Caco-2 Cells After 24 H Incubation, Assessed by MTT Assay. The FBS Control (Cells Without Hydrolysates) Was Defined as 100% Viability, And Treatment Values Are Shown Relative to This Control. Distinct Letters (A, B, C) Denote Significant Differences Between Means (P < 0.05).
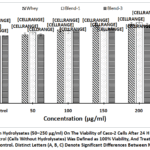 |
Figure 1: Effect of Protein Hydrolysates (50–250 µg/ml) On The Viability of Caco-2 Cells After 24 H Incubation, Assessed by MTT Assay. The FBS Control (Cells Without Hydrolysates) Was Defined as 100% Viability, |
Bioavailability of Plant Protein Blends
The proportion of essential and non-essential amino acids accessible for absorption from whey and plant protein blends was assessed using the Caco-2 cell monolayer model. After a 2-hour incubation, PlPI-blend 1 protein demonstrated the highest essential amino acid (EAA) transport (25%), followed by whey (16%) and PlPI-blend 2 (15%) (Table 3). Similarly, the transport of non-essential amino acids (NEAAs) was highest in the PlPI-blend 1 (62%), compared to 57% in whey protein and 40 % in PlPI-blend 2. Amino acid profiling revealed distinct transport patterns across the intestinal epithelial (Caco-2) barrier, reflecting the unique absorption characteristics of each protein source (Fig. 2).
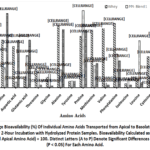 |
Figure 2: Percentage Bioavailability (%) Of Individual Amino Acids Transported from Apical to Basolateral Compartments in Caco-2 Cells After 2-Hour Incubation with Hydrolyzed Protein Samples. |
PlPI-blend 1 showed higher transport of neutral and polar amino acids, such as leucine (59.1%), isoleucine (10.6%), and proline (96.8%). While PlPI-blend 2 exhibited higher uptake of charged amino acids, including serine (93.2%), glutamic acid (59.8%), histidine (65.6%), lysine (56.5%), and aspartic acid (59.5%). Sulfur-containing amino acids, methionine and cysteine, were poorly absorbed in both plant blends.Whey showed superior bioavailability of critical EAAs like valine (74.4%) and phenylalanine (80.5%), higher than PlPI-blend 1 (6.3%) and PlPI-blend 2 (3.1). Conversely, PlPI-blend 2 exhibited the highest NEAA bioavailability of glutamic acid, arginine, serine and aspartic acid which are linked in literature to potential functional benefits for metabolic and cardiovascular health.However, direct bioactivity evaluations are required to confirm functional benefits. These results emphasize whey’s superior efficiency in delivering essential amino acids, while also highlighting the distinctive non-essential amino acid profiles and transport dynamics of plant protein blends.
Table 3: Absolute Concentrations of EAA and NEAA (µg/Gram of Protein ± SE) In (A) Apical and (B) Basal Compartments of Caco-2 Transwell Inserts
|
|
Upper phase | Lower phase | ||||
| Whey | PlPI- Blend 1 | PlPI- Blend 2 | Whey | PlPI- Blend 1 |
PlPI- Blend 2 |
|
|
Essential Amino Acids (µg/g protein) |
||||||
|
Threonine |
52.2±2 | 18.7±0.73 | 3.4±0.14 | 11.8±0.46 | 3.6±0.14 |
0.1±0.004 |
| Methionine | 2.2±0.09 | 2.1±0.08 | 7.4±0.29 | 1.5±0.06 | 0.6±0.02 |
2.3±0.09 |
|
Valine |
0.2±0.01 | 15.9±0.62 | 16.4±0.64 | 0.1±0.01 | 1±0.04 | 0.5±0.02 |
| Phenylalanine | 1.8±0.07 | 16.8±0.66 | 1.9±0.07 | 1.4±0.06 | 6.9±0.27 |
0.6±0.02 |
|
Isoleucine |
39±1.52 | 29.2±1.14 | 31.6±1.23 | 2.1±0.08 | 3±0.12 | 1.2±0.05 |
| Leucine | 2.8±0.11 | 4.4±0.17 | 2.2±0.09 | 0.3±0.01 | 2.6±0.1 |
0.1±0.01 |
|
Histidine |
0.4±0.02 | 8.2±0.32 | 6±0.23 | 0.3±0.01 | 4.3±0.17 | 3.9±0.15 |
| Lysine | 32±1.25 | 33.6±1.31 | 3.4±0.13 | 3.6±0.14 | 10.1±0.39 |
1.9±0.07 |
|
∑EAA |
130.75 | 128.95 | 72.52 | 21.27 | 32.10 | 10.76 |
|
Non-Essential Amino Acids (µg/g protein) |
||||||
| Arginine | 1±0.04 | 15.5±0.61 | 3.7±0.14 | 0.8±0.03 | 3.2±0.12 |
1.5±0.06 |
|
Serine |
0.4±0.02 | 15.3±0.6 | 0.3±0.01 | 0.2±0.01 | 3.6±0.14 | 0.3±0.01 |
| Aspartic acid | 0.7±0.03 | 66.8±2.61 | 3.2±0.12 | 0.4±0.01 | 4±0.16 |
1.9±0.07 |
|
Glutamic acid |
36.1±1.41 | 8.7±0.34 | 2.4±0.09 | 4.6±0.18 | 3.4±0.13 | 1.4±0.05 |
| Glycine | 13.7±0.53 | 12.6±0.49 | 2.1±0.08 | 1.9±0.07 | 2.2±0.08 |
0.1±0.002 |
|
Alanine |
0.3±0.01 | 551.7± 21.52 | 0.4±0.01 | 0.2±0.01 | 123.7±4.82 | 0.3±0.01 |
| Tyrosine | 45.1±1.76 | 3.3±0.13 | 3.1±0.12 | 3.8±0.15 | 0.3±0.01 |
0.3±0.01 |
|
Proline |
548.6±21.4 | 820.6±32 | 750.5±29.27 | 353.8±13.8 | 791.7±30.88 | 536.6±20.93 |
| Cysteine | 0.8±0.03 | 6.6±0.26 | 616.1±24.03 | 0.3±0.01 | 0.6±0.03 |
4.5±0.18 |
|
∑NEAA |
646.7231 | 1501.111 | 1381.665 | 365.8444 | 932.637 |
546.7709 |
Gene Expression Analysis
PepT1 (SLC15A1), a proton-coupled peptide transporter positioned on the apical surface of enterocytes, plays a pivotal role in di- and tripeptide absorption, accounting for approximately 80% of total dietary protein uptake.SREBF2 acts as a central transcriptional regulator of cholesterol balance by controlling the activity of genes linked to cholesterol production and absorption. To investigate the relationship between amino acid bioavailability and transporter activity, gene expression analysis was performed on Caco-2 cells exposed to protein hydrolysates. Whey protein hydrolysate maximally induced the upregulation and down regulation of PepT1 expression (1.47-fold) and SREBF2 (0.29-fold), a key regulator of cholesterol biosynthesis respectively. In comparison, PlPI-blend 1 and PlPI-blend 2 moderately elevated PepT1 expression (1.34-fold and 1.31-fold, respectively) and suppressed SREBF2 expression to 0.62-fold and 0.61-fold, respectively (Fig. 3). These results suggest that while plant protein blends can modulate nutrient transport and cholesterol-regulating gene expression, whey protein remains more effective in enhancing peptide transport and downregulating lipid synthesis pathways due to its higher essential amino acid bioavailability.
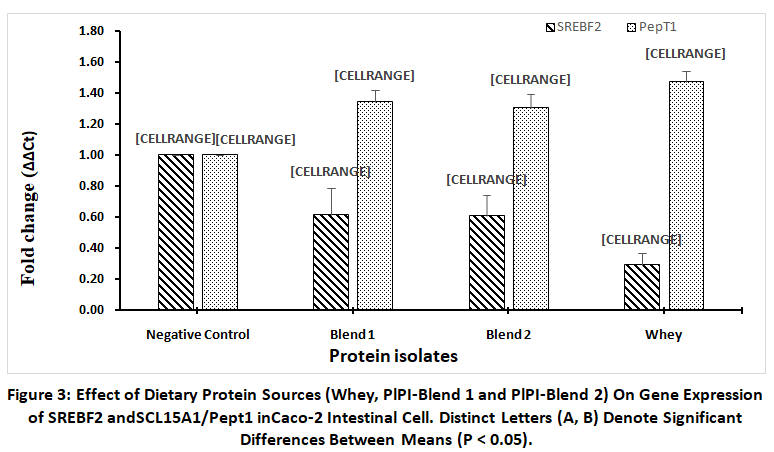 |
Figure 3: Effect of Dietary Protein Sources (Whey, PlPI-Blend 1 and PlPI-Blend 2) On Gene Expression ofSREBF2 andSCL15A1/Pept1 inCaco-2 Intestinal Cell. |
Discussion
This study demonstrated distinct nutritional and functional characteristics of whey protein isolate (WPI) and two formulated plant protein isolate blends (PlPI-blend 1 and PlPI-blend 2). While WPI exhibited the highest overall protein content, the PlPI blends showed comparable protein concentrations and greater solubility, with PlPI-blend 2 showing the highest soluble protein fraction. This higher solubility may enhance digestibility and functional utility in food applications.14
Cytotoxicity assays confirmed that all hydrolysed protein samples were non-toxic up to 250 μg/mL, a concentration below peak physiological levels in the intestine (~15,000 μg/mL equivalents post-meal), but representative of diluted in vitro conditions for safety assessment, supporting their safety for nutritional use. The observed enhancement in Caco-2 cell proliferation, particularly with PlPI-blend 2, indicates the release of bioactive peptides during enzymatic hydrolysis that may contribute to gut health and tissue regeneration. These results align with previous findings from hydrolysates of chickpea and hemp seed, which also stimulated intestinal cell growth.16,17
This study assessed amino acid bioavailability and and their relevance to PepT1-mediated transport (oligopeptides + free amino acid) using a fully differentiated Caco-2 in vitro model, comparing hydrolysed WPI to two PlPI-blends. Amino acid transport was measured as absolute concentrations (μg/g protein) in apical and basolateral compartments, reflecting the quantity absorbed relative to initial luminal amounts. Amino acid bioavailability studies revealed superior essential amino acid (EAA) bioavailability in WPI, particularly for branched-chain and sulfur-containing amino acids critical for muscle synthesis and metabolic regulation. In contrast, PlPI-blends, especially PlPI-blend 2, demonstrated enhanced transport of NEAAs such as aspartic acid, arginine, and glutamic acid, suggesting complementary metabolic benefits. Recent research in 2024 has shown that PlPIs often contain reduced levels of EAAs, which may diminish their effectiveness in promoting muscle protein synthesis when compared to whey proteins. Blending various plant proteins can enhance the overall EAA composition (as seen in PlPI-blend 1) and more effectively support muscle synthesis.18Despite WPI’s overall advantage, PlPI-blend 1 exhibited markedly high leucine bioavailability (47 % more that of whey), which may support mTOR signalling and muscle protein synthesis,potentially benefiting populations with high anabolic demands, including the elderly or plant-based dieters.19
Differences in digestibility, and anti-nutritional factors as given in Table 2 likely contribute to the varied bioavailability pattern observed between plant and animal proteins (Fig.2; Table 3).20,21Additionally, the alkaline extraction method used for PlPI-blends may lead to racemization of certain NEAAs, further limiting EAA bioavailability.22 Non-essential amino acids (NEAAs) such as D-enantiomers of aspartic acid, alanine, and glutamic acid are particularly prone to racemization during alkaline processing, potentially impairing the bioavailability of essential amino acids (EAAs).23,24 In contrast, whey protein, which contains a higher proportion of EAAs relative to NEAAs and undergoes less aggressive processing, is less susceptible to racemization. This likely contributes to the superior digestibility and amino acid bioavailability observed in whey protein isolate in this study. Supporting evidence from previous research also indicates that, although wheat protein hydrolysate can increase circulating amino acid levels, whey protein produces a more pronounced postprandial rise in blood EAA concentrations, reinforcing its efficiency in essential nutrient delivery.25Furthermore, recent 2024 study have reported that PlPIs generally have lower contents of EAA like methionine in comparison to animal protein, consistentwith our observations.26
Gene expression analysis corroborated these findings. WPI induced the highest upregulation of PepT1, a key intestinal peptide transporter, and the greatest suppression of SREBF2, a regulator of cholesterol biosynthesis.27While PlPI blends moderately influenced both genes, the data suggest that WPI more effectively facilitates amino acid absorption and lipid metabolism regulation.Our study demonstrated that whey protein hydrolysate significantly downregulated the expression of SREBF2, a key transcription factor involved in cholesterol biosynthesis, compared to more modest reductions observed with PlPI-blend 1 and PlPI-blend 2. This suggests a greater potential for whey protein to suppress endogenous cholesterol synthesis. Additionally, all protein treatments upregulated the expression of PepT1 (SLC15A1), a peptide transporter essential for di- and tripeptide absorption in the intestine. These findings align with existing studies demonstrating that high-quality protein diets enhance PepT1 expression, likely driven by increased availability of bioactive peptides and amino acids released during digestion. For example, in a recent 2025 study, ultrasonicated pea protein hydrolysates produced inhibitory peptides like SPR and YPR, which were transported via PepT1 but degraded by brush border peptidases, while others like YPHYR (bioactive peptide) remained partially intact and were ATP-dependent transported. These findings highlight how plant proteins can be important sources of bioactive peptides capable of modulating metabolic pathways.28Similar trends have been reported in animal models, where dietary supplementation with protein hydrolysates elevated PepT1 expression in the intestinal tissues of fish and poultry, further supporting the regulatory role of protein quality in nutrient absorption and cholesterol metabolism.29,30
These results highlight the role of protein-derived peptides in modulating cholesterol-related pathways, potentially via mTOR and MAPK signalling, as well as interaction with cholesterol transporters such as ABCA1. Transporters such as SLC38A9 (arginine transporter) and SLC6A15 (leucine and branched-chain amino acid transporter) mediate amino acid influx across the plasma membrane, initiating downstream signaling. In conditions of amino acid scarcity, sensors like CASTOR1 and Sestrin bind to and inhibit GATOR2, a positive regulator of mTORC1. However, upon amino acid sufficiency, these sensors dissociate, permitting GATOR2 to activate GATOR1, a GTPase-activating protein, which ultimately facilitatesthe enlistment of mTORC1 to the lysosomal membrane and its subsequent activation.31Activated mTORC1 enhances protein synthesis and upregulates amino acid transporter genes via ATF4-dependent transcription, consistent with our finding of increased SLC15A1 (PepT1) expression, which likely augments peptide and amino acid uptake.32 Furthermore, hyperactivation of mTORC1 initiates a feedback mechanism that suppresses Akt phosphorylation, thereby reducing phosphorylation of Insig proteins.33 Unphosphorylated Insig remains bound to SCAP, preventing cleavage and activation of SREBPs. As a result, SREBP2 remains sequestered in the endoplasmic reticulum, signaling cellular cholesterol sufficiency and leading to the observed downregulation of SREBF2.34 These interconnected regulatory networks provide a plausible mechanistic basis for the differential gene expression patterns observed in our protein-treated Caco-2 cells.Taken together, our findings indicate that the superior bioavailability and regulatory effects of WPI arise from its higher EAA content, minimal racemization during processing, and the presence of bioactive peptides capable of stimulating intestinal transporters like PepT1. This enhanced peptide uptake likely facilitates rapid postprandial increases in plasma amino acid levels, which in turn may activate mTOR and MAPK signaling pathways that influence lipid metabolism, as reflected by reduced SREBF2 expression and potential modulation of cholesterol synthesis. In contrast, PlPI blends show lower bioavailability, likely due to anti-nutritional factors and racemization of NEAAs during alkaline extraction, which may disrupt peptide structure, reduce transporter affinity, and impair absorption kinetics. These compositional and processing differences between animal and plant-derived protein isolates appear to translate directly into differential gene expression patterns and metabolic responses, highlighting the critical interplay between protein source, processing methods, and downstream physiological effects. A limitation of our work is the reliance on in vitro Caco-2 models, which, while informative, may not capture the full complexity of human digestion and metabolism. While peptides are generally more functional than free amino acids due to their bioactive sequences, this study did not characterize peptide hydrophobicity or size. Future studies should employ additional analyses, such as RP-HPLC and SEC, could evaluate peptide profiles to better link structure to function.Collectively, the findings support the use of PlPI blends as functional dietary components have promising metabolic benefits. However, WPI remains superior in EAA delivery and gene regulatory impact, reinforcing the potential of combining plant and animal proteins to optimize health outcomes. At this point, we have primarily focused on evaluating the digestibility, amino acid bioavailability, and cytotoxicity of the PlPI blends. Additional research, including in vivo experiments, protein level validation and metabolomics analyses, are needed to elucidate the mechanisms governing amino acid absorption and utilization, to comprehensively understand the bioavailability and physiological impacts of plant-derived protein isolates, and to explore whether combining plant and animal proteins could synergistically optimize both amino acid delivery and lipid metabolic outcomes.
Conclusion
This study compared two plant protein isolate blends (PlPI-blend 1 and 2) with whey protein isolate (WPI), revealing that WPI had higher essential amino acid bioavailability and stronger gene modulation, while PlPI-blends demonstrated superior solubility, cell-supportive activity, and non-essential amino acid uptake. These findings point to the complementary strengths of whey and plant proteins, emphasizing the potential of tailored protein blends to meet diverse nutritional, functional health needs that supports sustainable strategies for muscle health and cholesterol regulation. The conclusions are limited by the in vitro nature of the study and the comparatively lower bioavailability of EAA in PlPI blends hydrolysates. Future in vivo investigations are needed to fully assess the nutritional implications of these blends in real-world dietary contexts.
Acknowledgement
We would like to thank the Department of Agricultural Biotechnology at theNational Agri-Food Biotechnology Institute, India, for providing laboratory facilities.
Funding Sources
The authors acknowledge Indian Council of Agricultural Research-Indian Agricultural Research Institute (IARI), Department of Biotechnology (Project Sanction no. DBT BioE3 BT/PR59742/FFSP/171/18/2025) and Department of Science and Technology (DST) (Project Sanction no. DST/SHRI/MP/2023/51; IARI code: DST/SHRI/24-833/TG3714) for financial support.
Conflict of Interest
The author(s) do not have any conflict of interest.
Data Availability Statement
The manuscript incorporates all datasets produced or examined throughout this research study.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to Reproduce Material from Other Sources
Not Applicable
Author Contributions
- Brijesh Lekhak: Writing Original Draft, Methodology, Investigation, Data Curation, Formal Analysis.
- Minakshi Dutta: Investigation, Data Curation.
- Nitin Kumar Singhal: Methodology, Resources, Validation.
- Rama Prashat Govindarajalu: Statistical Analysis, Resources
- Navita Bansal: Methodology.
- Suneha Goswami: Methodology, Validation.
- Ranjeet Ranjan Kumar: Methodology.
- Aruna Tyagi: Conceptualization, Methodology, Supervision, Validation, Review and Editing.
- Vinutha Thimmegowda: Conceptualization, Funding Acquisition, Methodology, Validation, Visualization, Data Curation, Review and Editing.
References
- Vallejo-Vaz AJ, Ray KK. Epidemiology of familial hypercholesterolaemia: community and clinical. Atherosclerosis. 2018;277:289-297.
CrossRef - Shahnaz T, Fawole AO, Adeyanju AA, et al. Food proteins as functional ingredients in the management of chronic diseases: a concise review. Nutrients. 2024;16(14):2323. doi:10.3390/nu16142323
CrossRef - Mei L, Fu Q, Guo T, et al. Structural changes and cholesterol-lowering in denatured whey protein isolate: malic acid combined enzymolysis. Food Hydrocolloids. 2022;127:107502. doi:10.1016/j.foodhyd.2022.107502
CrossRef - Jenkins DJ, Kendall CW, Faulkner DA, et al. Assessment of the longer-term effects of a dietary portfolio of cholesterol-lowering foods in hypercholesterolemia. Am J Clin Nutr. 2006;83(3):582-591.
CrossRef - El-Adawy TA, Taha KM. Characteristics and composition of different seed oils and flours. Food Chem. 2001;74(1):47-54.
CrossRef - Kaul P. Nutritional potential, bioaccessibility of minerals and functionality of watermelon (Citrullus vulgaris L) seeds. LWT Food Sci Technol. 2011;44(8):1821-1826.
CrossRef - Olivos-Lugo BL, Valdivia-López MÁ, Tecante A. Thermal and physicochemical properties and nutritional value of the protein fraction of Mexican chia seed (Salvia hispanica L). Food Sci Technol Int. 2010;16(1):89-96.
CrossRef - Lakshmi YD, Kumar RD, Dutta M, et al. Improved nutritional and functional properties of plant protein isolate blends through steam infusion. Food Chem. 2025;464:141863. doi:10.1016/j.foodchem.2024.141863
CrossRef - Du M, Xie J, Gong B, et al. Extraction, physicochemical characteristics and functional properties of mung bean protein. Food Hydrocolloids. 2018;76:131-140.
CrossRef - Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248-254.
CrossRef - Vinutha T, Kumar D, Bansal N, et al. Thermal treatments reduce rancidity and modulate structural and digestive properties of starch in pearl millet flour. Int J Biol Macromol. 2022;195:207-216.
CrossRef - Biswal BK, El SM, Kumari D, et al. Twin function of zein–zinc coordination complex: wheat nutrient enrichment and nanoshield against pathogenic infection. ACS Sustain Chem Eng. 2018;6(5):5877-5887.
CrossRef - Goulart AJ, Bassan JC, Barbosa OA, et al. Transport of amino acids from milk whey by Caco-2 cell monolayer after hydrolytic action of gastrointestinal enzymes. Food Res Int. 2014;63:62-70.
CrossRef - Chaudhary R, Singh S, Kaur K, et al. Genome-wide identification and expression profiling of WOX genes in banana. 3 Biotech. 2022;12(11):321. doi:10.1007/s13205-022-03387-w
CrossRef - Dai Y, Lu X, Li R, et al. Effects of ultrasound treatment on structure, functional properties and in vitro digestion of Sipunculus nudus protein. Int J Biol Macromol. 2024;277:134422. doi:10.1016/j.ijbiomac.2024.134422
CrossRef - Girón-Calle J, Alaiz M, Vioque J. Effect of chickpea protein hydrolysates on cell proliferation and bioavailability. Food Res Int. 2010;43(5):1365-1370.
CrossRef - Logarušić M, Slivac I, Radošević K, et al. Hempseed protein hydrolysates’ effects on proliferation and oxidative stress. Mol Biol Rep. 2019;46:6079-6085.
CrossRef - Lim C, Janssen TA, Currier BS, et al. Muscle protein synthesis in response to plant-based protein isolates versus whey protein. Curr Dev Nutr. 2024;8(6):103769. doi:10.1016/j.cdnut.2024.103769
CrossRef - Cui C, Wu C, Wang J, et al. Leucine supplementation alters placental metabolism via PI3K/AKT/mTOR pathway. Food Funct. 2022;13(4):2083-2097.
CrossRef - Bos C, Gaudichon C, Pueyo ME, et al. Postprandial kinetics of dietary amino acids after soy or milk protein ingestion. J Nutr. 2003;133(5):1308-1315.
CrossRef - Devappa RK, Swamylingappa B. Biochemical and nutritional evaluation of Jatropha protein isolate. J Sci Food Agric. 2008;88(5):911-919.
CrossRef - Gilani GS, Xiao CW, Cockell KA. Impact of antinutritional factors on protein digestibility. Br J Nutr. 2012;108(suppl 2):S315-S332.
CrossRef - Csapó J, Albert C, Csapóné KZ. The D-amino acid content of foodstuffs: a review. Acta Univ Sapientiae Alimentaria. 2009;2:5-30.
- Genchi G. An overview on D-amino acids. Amino Acids. 2017;49:1521-1533.
CrossRef - Gorissen SH, Horstman AM, Franssen R, et al. Ingestion of wheat protein increases muscle protein synthesis in older men. J Nutr. 2016;146(9):1651-1659.
CrossRef - Auer J, Alminger M, Marinea M, et al. Digestibility and bioavailability of plant-based proteins and minerals. LWT. 2024;197:115893. doi:10.1016/j.lwt.2024.115893
CrossRef - Brown MS, Goldstein JL. The SREBP pathway: regulation of cholesterol metabolism. Cell. 1997;89(3):331-340.
CrossRef - Zhao M, Lei J, Yan Q, et al. Discovery of pea protein-derived DPP-IV inhibitory peptides. Int J Biol Macromol. 2025;307:141897. doi:10.1016/j.ijbiomac.2025.141897
CrossRef - Bakke S, Jordal AEO, Gómez-Requeni P, et al. Dietary protein hydrolysates affect PepT1 expression in cod. Comp Biochem Physiol B Biochem Mol Biol. 2010;156(1):48-55.
CrossRef - Gilbert ER, Li H, Emmerson DA, et al. Dietary protein quality influences nutrient transporter mRNA. J Nutr. 2008;138(2):262-271.
CrossRef - Chantranupong L, Scaria SM, Saxton RA, et al. CASTOR proteins are arginine sensors for mTORC1 pathway. Cell. 2016;165(1):153-164.
CrossRef - Park Y, Reyna-Neyra A, Philippe L, et al. mTORC1 balances amino acid supply via ATF4. Cell Rep. 2017;19(6):1083-1090.
CrossRef - Yoon MS. The role of mammalian target of rapamycin in insulin signaling. Nutrients. 2017;9(11):1176.
CrossRef - Xiao X, Tontonoz P. PUFAs regulate SREBP1c through Insig2 phosphorylation. Proc Natl Acad Sci U S A. 2024;121(40):e2416363121. doi:10.1073/pnas.2416363121
CrossRef
Accepted on: 30 Sep 2025
Second Review by: Yuosra Amer Ali
Final Approval by: Dr. Mattia Di Nunzio





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








