Characterization of Polyphenol Oxidase (PPO) from Blackberry Thorny Wild Rubus Fruticosus and its Inhibition using Natural Extracts
 , , 4, Mohammed Reda Britel2, , 3*
, , 4, Mohammed Reda Britel2, , 3* 1Biotechnology and engineering of biomolecules equip, Faculty of Sciences and Techniques, Abdelmalek Essâadi University, Tetouan, Morocco.
22Laboratory of Technology and Innovation, National School of Applied Sciences of Tangier, University Abdelmalek Essaadi, Tetouan, Morocco.
3Faculty of Sciences and Techniques, University Abdelmalek Essaadi, Tetouan, Morocco.
4Engineering, Innovation, and Management of Industrial Systems Laboratory, Faculty of Sciences and Techniques, University Abdelmalek Essaadi,
Corresponding Author Email: amal.maurady.ma@gmail.com
DOI : http://dx.doi.org/10.12944/CRNFSJ.10.3.33
Download this article as:
![]()
The Polyphenol oxidase (PPO) leads to the enzymatic browning of fruits and vegetables and needs to be prevented in food browning and quality. The present study aimed to investigate the use of natural extracts and chemical inhibitors to prevent browning of the PPO of blackberries. Purification, characterization, and kinetics of PPO of blackberry parameters for five substrates, namely, pyrocatechol, 4-methylcatechol, Pyrogallol, Gallic acid, and tyrosine, were described. The results showed that the DEAE-Sepharose and Superdex G-200 purification methods, which achieved electrophoretic purity, increased PPO activity by 556 fold. Purification with Sephadex GE-200 and SDS-PAGE reveals two PPO isoenzymes with an apparent molecular weight of 22 kD and 70 kD. The optimum pH and temperature values indicated were 6.6 and 25°C, respectively. The PPO showed variable affinity towards o-dihydroxy phenolic substrates with catecholase activity but without any activity observed with phenol, a monohydroxy substrate, and it was very effective towards pyrocatechol, pyrogallol, and 4-methyl catechol. The results revealed that inhibition of the PPO using both synthetic inhibitors and natural extracts was the most effective method. Quercetin and ascorbic acid showed higher inhibition with the lowest Ki values. Fresh onion (Allium cepa) and wild Arbutus unedo extract were able to inhibit the blackberry PPO activity up to 50% and 60%, respectively. Therefore, the use of natural extracts from Arbutus unedo L as anti-browning agents on the blackberry PPO may provide new insight to overcome the enzymatic browning.
KEYWORDS:Arbutus Unedo L; Anti-Browning Agents; Natural Extracts; Polyphenol Oxidase
Introduction
The thorny wild blackberry (Rubus fruticosus) has a short post-harvest lifespan; therefore, it loses its quality due to its extremely fragile structure and breathing high rate. It cannot be kept in the air, even for a short period of two or three days 1. The consumption of this fruit is still considered very low 2-3 despite being abundant in nutritious components, such as: vitamin C, bioactive chemicals, and functional molecules with biological activities. Blackberry is rich in antioxidants that play an important role in our health as it contains polyphenol, which is one of the most important antioxidants. This phenolic is highly oxidized by polyphenol oxidase (PPO) (monophenol, dihydroxyphenylalanine) (oxygen oxidoreductase; E.C. 1.14.18.1.), to its corresponding quinone. The quinone produced is a highly reactive substance that normally reacts further with other quinones, amino acids or proteins to produce coloured compounds responsible for food quality deterioration 4-5.The organoleptic quality may change adversely due to the enzymatic oxidation of phenolic compounds. PPO has been studied in a wide variety of fruits. In the presence of oxygen, PPO catalyzes two types of reactions: hydroxylation of monophenols to o-diphenols and oxidation of o-diphenols to o-quinones, representing monophenolase (cresolase) and diphenolase (catecholase) activity, respectively 6,7,8. PPO is a major contributor to enzymatic browning or color degradation in fruits and vegetables during postharvest processing operations due to these reactions 9-10.
The PPO is vital for wound healing, pathogen defense, and many cellular processes, such as oxygen level control in chloroplasts 6-11. As reported by Yoruk 5, browning reaction affected the sensory quality with diminution marketing of fruit by lowering nutritional quality.Quieroz et al. 12 also suggested that the enzymatic browning leads not only to color alteration and antioxidant degradation but also organoleptic and nutritional losses because of the quinones’ condensation with other compounds such as amino acids, proteins, phenols, and sugar.
In food industry, the inhibition of PPO enzyme and prevention of browning is a big challenge for fruit and vegetable 6. Most of the anti-browning systems, both physical as well as chemical methods, have been used to inhibit PPO activity and thereby prevent browning 13. Thermal technologies are the most commonly used physical-based preservation systems for processing juice 14.
However, one of the study reported a negative impact of heat processing on the organoleptic and nutritional qualities of final products 15. Chemical-based methods to control enzymatic browning in postharvest are related to the application of PPO inhibitors such as sulphur, cysteine ascorbic acid, citric acid cyclodextrins, and oxalic acid 16. However, those synthetic additives have a potential hazard on human health 17,18.
Browning effect can occur within a short time leading to deterioration of nutritional value and appearance of Blackberry 19. The characterization studies of PPO of the blackberry were reported in a previous study 19 without inhibition and purification. These data are very important during the processing of the wild blackberry, rich in antioxidants and anthocyanin. Natural extract has been studied for PPO inhibitors for potato 20, pear 21 and banana 22. However, the study of anti-browning agents’ effects on blackberry needs to be established. In this research, we try to reveal more information on the PPO purification, chemical and natural anti-browning agents in blackberry, which is yet to be established. There is no scientific information on how the natural and chemical inhibitors may influence the PPO activity of blackberries. Thus, the inhibitory effects of chemical and natural inhibitors (onion and Arbutus unedo extracts) with their kinetics on blackberry PPO were thus investigated in this work.
The objective was to report all the properties that have never been mentioned before, regarding the purification of PPO, effects of pH and temperature on PPO activity and substrate specificity, performed with pyrocatechol, 4-methylcatechol, pyrogallol, gallic acid, and tyrosine. Inhibition study of the blackberry thorny wild PPO using natural and chemical inhibitors was investigated to show the inhibitory effects on our enzyme. These data may be used in testing effective methods to inhibit discoloration naturally without adversely affecting our health, while avoiding the breakdown of anthocyanins in blackberry juice and other processed products.
Materials and Methods
Plant material and natural inhibitors
Blackberry thorny wild (Rubus fruticosus) originated from local coast (Slookia (35°47’16.3″N 5°54’31.3″W) Tangier, Morocco, was used in this study. Red onion (Allium cepa) and Wild Arbutus unedo L, were local products also. All chemicals used in this study were analytical grade.
PPO Extraction
The extraction procedure of PPO from the blackberry thorny wild was performed according to Yagar [23]. 200 g of the blackberry thorny wild was homogenised with 200 mL of 0.1 M phosphate buffer, pH 6.8, containing 10 mM ascorbic acid, 0.1% polyvinylpyrrolidone and 0.5% Triton X-100 using a industrial blender (Waring Laboratory ScienceTM LB20EG, French) at 5,000 rpm for 3 min. The homogenate was filtered through filter paper and the filtrate was centrifuged at 160000 xg for 30 min at 4 °C by an ultracentrifuge (Centrifuge Universal 320R, Hettich Zentrifugen, Tuttlingen, Germany). The supernatant was used as the crude enzyme extract. Then it was brought to 30-90% (NH4)2SO4 saturation, pH 6.8. The fractions were solubilized in 30 mL of phosphate buffer (pH 6.8, 0.1 M) and dialyzed, using a dialysis cassette of 3,5 K cut off the membrane in 2000 mL of phosphate buffer (pH 6.8, 0.1M) at 4 °C, with three buffer changes at 6 and 8 hour intervals for salt removal.
Superdex gel filtration chromatography
The dialysed enzyme extract was chromatographed on a Sephadex 200 GE column according to Mishra [10]. The protein was loaded onto a column and the elution was carried out in Tris-HCL buffer (pH 8.0, 2 mM) with 150 mM of NaCl. The flow rate was maintained at 1 mL/min and the detection was carried out using an UV detector set at 280 nm. The PPO activity and protein estimation were also performed for different fractions collected at 30 s intervals.
DEAE anion-exchange chromatography
The final step of purification was conducted using DEAE. Sepharose column material (40 ml) was prepared according to the method of Mishra et al.10, with some modifications. Initially, washing was done with distilled water (150 mL) and then with Tris-HCl buffer (pH 8.10 mM). The fractions containing activity were pooled and subjected to anion exchange chromatography and mixed with DEAE column material, kept at slow stirring for 30 min at 4 °C. This slurry was then washed twice with 250 mL of Tris-HCL buffer (pH 8.0, 2 mM) and loaded onto the column. The elution was carried out using an increasing gradient of NaCl from 0 to 0.5 M in 150 mL of tris-HCl buffer (pH 8.0, 2 mM). The flow rate was 1 mL/min and fractions of 1 mL were collected. PPO activity was determined for each fraction.
Electrophoresis (SDS-PAGE)
Sodium dodecyl sulphate polyacrylamide gel electrophoresis under reducing and denaturing conditions (SDS–PAGE) (10%) was carried out in the same way as described by Davis and Laemmli 24 . The protein samples were prepared by mixing 2× (double strength) gel loading buffer (0.1M Tris-C1, pH 6.8, 4% SDS, 0.2% bromophenol blue, 20% glycerol) and β-mercaptoethanol (0.2 M), mixed in a proportion of 1:1, boiled for 5 min, centrifuged (10,000g) for 2 min, and then loaded onto the gel, along with the molecular weight marker mix. We adjusted the electrophoresis to 25–40 mA with a duration of 4 h at room temperature. The quantity of the proteins deposited in the well was 20 μg. Following electrophoresis, the gel was fixed in acidified methanol water (40% methanol and 10% acetic acid) for 1 hour before being colored in 0.1% Coomassie brilliant for 3 hours on a rocker (Neolab, Mumbai, India). Later, the gel was de-stained with the same acidified methanol–water until the protein bands became distinguishable and prominent. The substrate staining of the gel was performed with pyrocatechol (0.05 M) substrate in phosphate buffer (pH 6.6, 0.2 mM) for up to 1 h, until the bands became prominent.
Assay of protein and polyphenol oxidase (PPO) activity
The enzyme activity was assayed according to the method of Gonzalez [19] with some modification. The reaction mixture consisted of 2.5 mL of 0.05 M pyrocatechol in 0.2 M phosphate buffer (pH 6.5) and 0.5 mL of PPO extract. The increase in absorbance at 420 nm was monitored at 30 s intervals for 3 min using a spectrophotometer (Model UV-3100), and the average change in absorbance per min was calculated. The reaction velocity (V) was calculated from the linear part of the plot of absorbance (A) against time (t). The unit of PPO activity was defined as the change in the absorbance of 0.001 min (DA 420 nm/min) due to the oxidation of the substrate. The PPO activity was expressed as U/g of blackberry weight. The specific activity was determined by expressing PPO activity/mg protein. The protein content of blackberry extract was determined by the Bradford method [25], using bovine serum albumin (BSA, Sigma Chemical, St. Louis, USA) as a standard. The total enzyme molecule concentration was determined by dividing the estimated total protein by the molecular weight of PPO. The catalytic efficiency (vmax/km) was calculated by dividing Vmax by Km.
Determination of substrate specificity
In order to determine the Michaelis constant (Km) and maximum velocity (Vm), PPO activities were measured using pyrocatechol, 4-methyl-catechol, pyrogallol, acid gallic and tyrosine. All compounds were prepared in a 0.2 M sodium phosphate buffer, pH 6.5. The changes in absorbance for substrates 4-methycatechol and pyrocatechol were monitored at 420 nm. The absorbances for pyrogallol, tyrosine, and gallic acid were monitored at 334, 472, and 350 nm, respectively. The absorbance was recorded at 30 s intervals for 3 min, and the average change in absorbance per min was calculated and termed as activity. The Km and Vmax values were calculated by plotting one/activity (1/V) versus one/substrate (1/[S]) using Lineweaver and Burk’s direct linear plot method 26. The relative activities of these substrates were expressed as percentages with respect to those of the pyrocatechol substrate (considered 100%).
pH and temperature profile of PPO
The optimum pH of the partially purified PPO was investigated by measuring its activity at room temperature (25 °C) at pH ranging from 5 to 8. The tests were carried out using 0.2 M sodium phosphate buffer. The effect of temperature on PPO activity was carried out by assaying the reaction mixture at various temperatures ranging from 10 ° C, 20 ° C, 30 ° C, 40 ° C, 50 °C, 60 °C, and 70 °C.
Inhibition studies
Effect of chemical anti-browning agents
The blackberry PPO was incubated in the presence of the following chemical compounds: reduced glutathione, ascorbic acid, L-cysteine, quercetin, sodium sulfate, citric acid, chloride acid, and sodium fluoride, to at 0.1, 1.0, and 10 mM.
Additionally, 50 mM pyrocathechol in 0.2 M phosphate buffer (pH 6.5) was added.The reaction mixture contained 2 mL of substrate, 0.5 mL of inhibitor solution, and 0.5 mL of enzyme. The percent inhibition was calculated using the following equation:
Inhibition (%) = (1-(Ai/A0)) × 100
Where A0 is the initial PPO activity (i, e. Without inhibitor), and Ai is the PPO activity with inhibitor.
The inhibitory concentration that reduced the enzyme activity by 50 % (IC50) was determined for the inhibitors with a lower percentage of PPO residual activity at a concentration of 0.1 mM using three different concentrations of inhibitor. Finally, PPO activities were measured at constant inhibitor concentrations (IC50) using five different substrate concentrations. The Lineweaver-Burk graphs were used to determine the type of inhibition.
Effect of natural anti-browning agents
Two natural sources, red onions and arboursier (Arbutus unedo), were prepared according to the Wong et al. 27 method. Thus, 100 g of each natural source was homogenized separately with 200 mL of 0.1 M phosphate buffer, pH 7.5, for 2 min using a waving blender (Waring Laboratory ScienceTM LB20EG, French) at maximum speed. The homogenized thus prepared from onion and arboursier mixture was centrifuged for 20 minutes at 4°C at 4,000 rpm. After centrifugation, the supernatant was filtered using filter paper, and the filtrate was collected.
The PPO activity was determined with and without the presence of natural anti-browning agents extracted from local fresh wild arboursier diluted fruit (6%) and fresh red onion. The reaction mixture contained 0.5 mL of the enzyme, 0.5 mL of 0.1 M phosphate buffer, pH 7.5, and 0.5 mL of fresh extract as an inhibitor, incubated for 5 min at 20°C. Then, 1.5 ml of pyrocatechol (0.05 M to 0.01 M) was added to the reaction mixture. The control contained 0.5 mL of PPO and 2 mL of substrate solution in 0.5 mL of 0.1 M phosphate buffer. The total volume of the assay was 3 ml. The linear graph of the absorbance versus time (in seconds) curve was plotted to determine the initial rate of PPO activity. The PPO activity obtained was used to calculate the percentage inhibition as compared to the initial PPO (Ao) activity without inhibitor 27.
Statistical analysis
All analytical values represent the meansand standard deviations were calculated of triplicate measurements. One way ANOVA (Significance threshold: P ≤ 0.05) was performed to ascertain the significance of the means. Pairwaise comparison were performed based on Tukey HSD test. Statistical analyzes were performed using the statistical software SAS (Statistical Analysis System. Version 9.1. 2002).
Results and discussion
Purification of PPO
Blackberry PPO was sequentially purified by ammonium sulfate with precipitation from 30% to 90% salt saturation, dialysis, chromatography to gel filtration, and anion exchange chromatography. The results are summarised in (Table 1). The purification resulted in about a 556 fold increase in enzyme activity, and the purified PPO had an overall activity yield of 3,13%, which is similar to the activity yield reported for PPO from banana 28, but with a different specific PPO activity of 315757,6 U/mg. The specific activity in crude extract was 568.2 U/mg protein, which is similar to the specific activity reported for PPO from Barbados cherry 29 and higher than that reported for PPO from Loquat 30.
During steps of purification, the PPO activity/mL increased and, as expected, the protein (PPO) content decreased. This resulted in an increase of specific activity of up to 556 fold after gel DEAE chromatography (Table 1). However, at each step of purification, a loss in total activity was noticed because only peak fractions displaying PPO activity were purposely collected. This significantly increased the purification-fold; however, the total yield was significantly reduced to about 3,13% (Table 1). PPO blackberry extract was dialyzed and chromatographed on Sephadex GE-200 (Figure 1 a). Fractions comprising two peaks are shown in (Figure 1a). The latter shows high activity of PPO towards pyrocatechol. The filtration gel separated our PPO according to their sizes, that is to say, into two different molecular weights The first eluent is the first peak, which corresponds to the large molecule of the PPO, and the second eluent corresponds to the smallest molecule of our PPO. Then these active fractions were pooled and subjected to anion-exchange chromatography (Figure 1b.) with Tris-HCL (2 mM, pH = 8) buffer.The bound enzyme was eluted with a linear gradient of NaCl (0–0.5 M). To test the activity, high protein fractions were collected.These fractions (peaks 20–26) corresponded to NaCl concentrations between 0.25–0.35M. The protein content of the elution was determined on a single peak (the second) using Coomassie Brilliant Blue and measured at 595 nm. Enzyme activity was determined using pyrocatechol and measured at 420 nm (Figure. 1 a). Active fractions, with high enzyme activities, were collected and pooled. The resulting pooled fractions were concentrated by ammonium sulfate precipitation. Finally, the precipitated protein was re-dissolved in a small volume of 0.1 M phosphate buffer (pH 6.8) and dialyzed with the same buffer to use it in the SDS gel.
Table 1: Purification of PPO from Blackberry.
| Purification step | Volume (mL) | Protein mg/mL | Protein total(mg) | Activity U/ml | Activity total(U) | Specific activity(U/mg) | Purification (fold) |
Yield % |
| Crude extract | 200 | 0,440 | 88 | 250 ±1,44 | 50000 | 568,2 | 1 | 100% |
| (NH4)2SO4 | 70 | 0,960 | 67,2 | 348 ± 1,20 | 24360 | 362,5 | 0,64 | 48% |
| Dialysate | 3 | 0, 300 | 0,9 | 460 ± 1,44 | 1380 | 1533,3 | 2,7 | 27,6% |
| (Sephadex 200 GE) | 2,5 | 0,055 | 0,138 | 950 ± 2,88 | 2375 | 17272,7 | 30,4 | 4,8% |
| Ion exchange | 1,5 | 0,003 | 0,005 | 1042 ±3,55 | 1563 | 315757,6 | 556 | 3,13% |
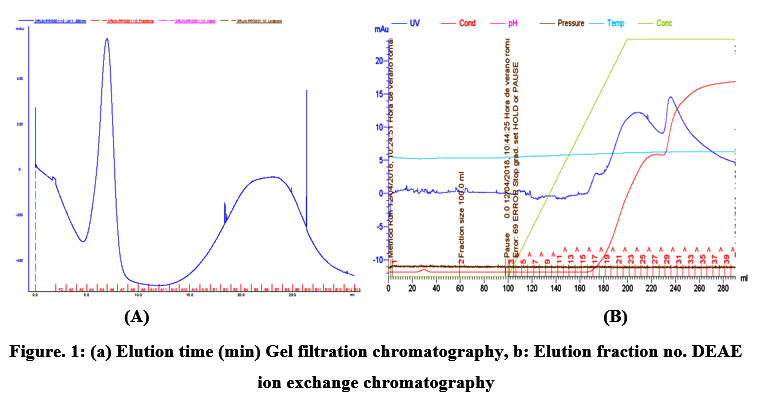 |
Figure 1: (a) Elution time (min) Gel filtration chromatography, 1b: Elution fraction no. DEAE ion exchange chromatography. |
PPO molecular weight
We have presented different purification steps on the SDS gel, and the amount of protein was minimal, which explains the appearance of thin strips. The crude extract showed us several bands which may be the bands of peroxidase and polyphenoloxidase, but during the following steps of purification of the PPO, two final bands clearly appear, one at 22 kD and the other at 70 kD at the level of the sephadex filtration gel, which proves the appearance of the two active peaks with different weights (Figure 2).
During the purification steps, the concentration of the enzyme was significantly reduced by DEAE, which proves the absence of the gel band. These results are similar to that of Barbados cherry 29, which also reveal two dominant bands but with molecular masses of 52 and38 kDa. Thus, this enzyme differs from PPO isolated from pear 31 and Chinese cabbage 32, and tomato PPO 33, which were reported to be monomers.
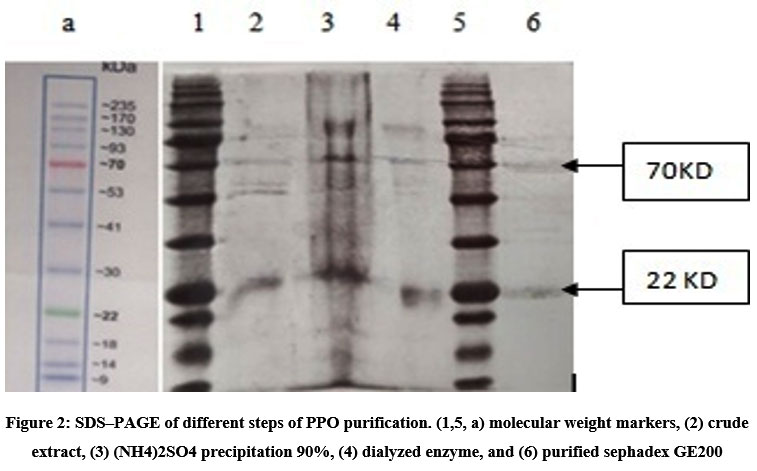 |
Figure 2: SDS–PAGE of different steps of PPO purification. (1,5, a) molecular weight markers, (2) crude extract, (3) (NH4)2SO4 precipitation 90%, (4) dialyzed enzyme, and (6) purified sephadex GE200 |
PPO substrate specificity
Five substrates, namely pyrocatechol, 4-methylcatechol, pyrogallol, gallic acid, and tyrosine, were tested and the PPO showed no activity using mono-hydroxyl phenol as substrates, low activity using tri-hydroxyl phenol, but displayed much greater activity with the o-dihydroxy phenol substrate (Table 2). Among the substrates tested, pyrocatechol had the maximum activity. The average enzymatic activity was observed even with the 4-methyl-catechol diphenolic substrate. On the other hand, the absence of the –CH3 group increased activity by 50%, as has been observed for pyrocatechol and is related to the role of the CH3 group and its location in the catechol structure, decreasing its specificity. These results are not similar to Bibhuti et al 34, as they found that the CH3 group increased its specificity. Trihydroxyphenol substrates such as pyrogallol and gallic acid exhibited low activity of the blackberry PPO. The monophenolic substrate, such as L-tyrosine, showed no activity against the blackberry PPO. These findings are similar to those reported for Yali pear peppermint, grapes Koshu PPO 35, and lonicera japonica thunb 36. These results revealed that the blackberry PPO was diphenolase, and there was no monophenolase or triphenolase activity.
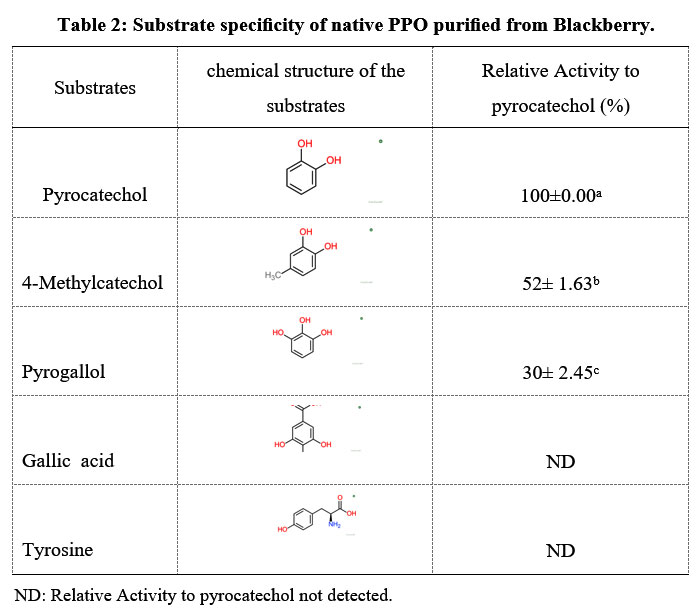 |
Table 2: Substrate specificity of native PPO purified from Blackberry. |
ND: Relative Activity to pyrocatechol not detected.
Kinetic characteristics of PPO
The Km and Vmax values were determined for substrates having higher activity (30%), using Lineweaver–Burk plots. Figures 3 to 5 show the Lineweaver-Burk curves of the mature PPO in the presence of the different substrates successively: pyrocatechol, 4-methylcatechol, and pyrogallol.
The best substrate for each enzyme depends on two factors: strong substrate binding or high affinity and high catalytic efficiency for a fixed enzyme concentration. The Vmax/Km ratio, referred to as “catalytic power”, can identify the most effective substrate.
The lowest Km was observed for pyrocatechol (0.0199 M). The 4-methyl catechol and pyrogallol showed higher Km values compared to pyrocatechol. The catalytic efficiency (Vmax/Km) was also found to be maximal for pyrocatechol and less for pyrogallol (Table 3). The values of Km and Vmax show that pyrocatechol is the best substrate for PPO in wild blackberry. These results are similar to several studies of lonicera japonica thunb (36), blackberry (19), and blueberry 37.
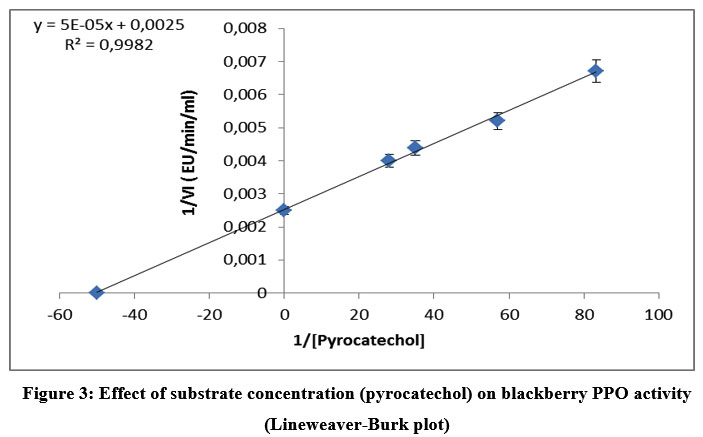 |
Figure 3: Effect of substrate concentration (pyrocatechol) on blackberry PPO activity (Lineweaver-Burk plot) |
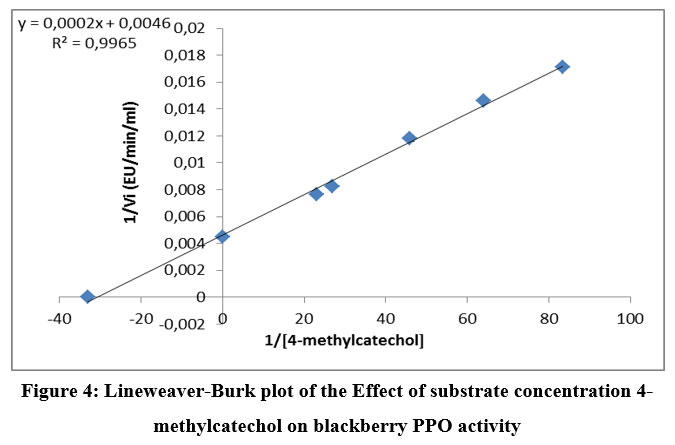 |
Figure 4: Lineweaver-Burk plot of the Effect of substrate concentration 4-methylcatechol on blackberry PPO activity |
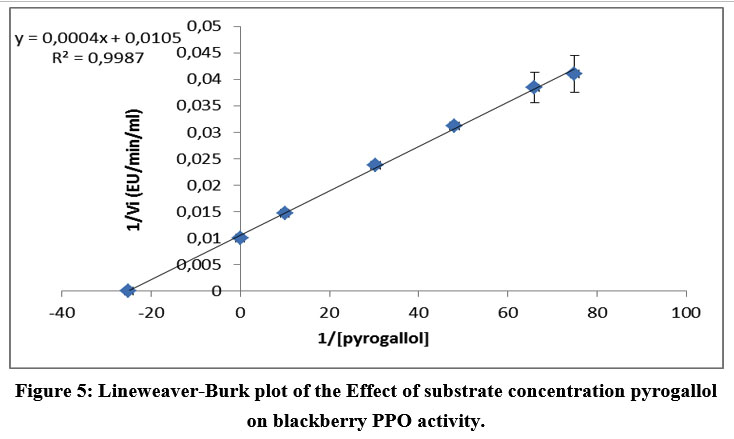 |
Figure 5: Lineweaver-Burk plot of the Effect of substrate concentration pyrogallol on blackberry PPO activity. |
Table 3: The kinetic characteristics of purified Blackberry PPO
| Substrates | Km (M) | Vmax (EU/min/mL) | Vmax/Km(EU/min. mL-1. M-1) |
| Pyrocatechol | 0.0199± 0.00b | 500± 20a | 25010± 17a |
| Pyrogallol | 0.039± 0.00a | 99± 13c | 2530± 27c |
| 4-Methyl catechol | 0.025± 0.00b | 250± 20b | 10014± 25 |
| Tyrosine | ND | ND | ND |
| GallicAcid | ND | ND | ND |
Km : Michaelis–Menten constant, Vmax : maximum reaction velocity, Vmax/Km : catalytic efficiency. The values are expressed as means ± SD of three parallel experiments. The values in column with different superscripts are significantly different (P ≤ 0.05).
ND: Not detected
Effect of temperature on the PPO activity
The blackberry PPO was active over a wide temperature range (10–70 °C), with the maximum activity being noted at 25 °C (Figure 6). Even at temperatures as high as 60 °C, the enzyme maintained 39% of its maximum activity; however, above this point, activity dropped dramatically to a minimum of 10% at 65 °C. The increase in the PPO activity as a function of temperature could be due to the fact that the increase in temperature enhances the kinetic energy, which leads to an acceleration of the reaction 38. However, at temperatures higher than 65 °C, we observed a reduction in the PPO activity, probably due to thermal denaturation of the enzyme.
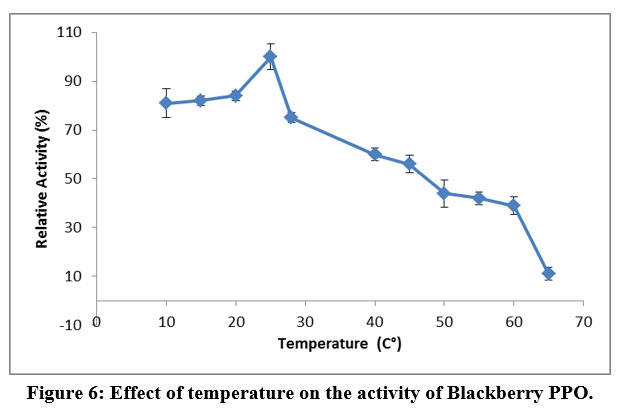 |
Figure 6: Effect of temperature on the activity of Blackberry PPO. |
Effect of pH
Blackberry PPO presented (Figure 7) maximum activity in the range of pH 6–7. Although the activity decreases below and above 6.6, the enzyme was still active at pH 5.4 with a remaining activity of 60 ∆OD/min/g. At pH 8.0, the remaining activity of the enzyme was 68 ∆OD/min/g. Several authors reported that the optimal pH for PPO activity varied from about 4.0 to 7.0 depending on the extraction methods, substrates, and localization of the enzyme in the cell 39.
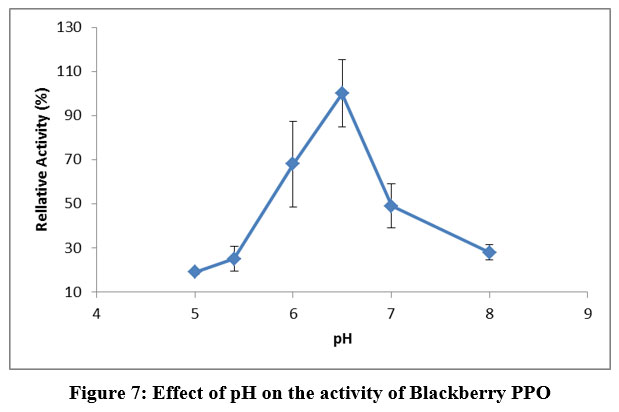 |
Figure 7: Effect of pH on the activity of Blackberry PPO. |
Inhibitor studies
Effect of chemical inhibitors on enzyme activity
The effect of various chemical antibrowning agents with different concentrations on the blackberry PPO was tested with pyrocatechol substrate (Table 4). The percentage inhibition increased with the increasing inhibitor concentrations. Ascorbic acid, reduced glutathione, and L-cysteine all strongly inhibited the enzyme, with PPO activity of 100%, 100%, and 93.33%, respectively, at a concentration of 10 mM, while quercitin could inhibit the enzyme at a low concentration of 0.1 mM with PPO activity of 89%. Furthermore, ascorbic acid, reduced glutathione, and quercetin have the lowest IC50 and Ki values among the other anti-browning agents, which were 0.18 mM, 0.2 mM, and 0.19 mM, respectively, except for L-cysteine, with 1.25 mM Ki. This further proves that L-cysteine, reduced glutathione, quercetin, and ascorbic acid have greater efficiency in inhibiting the blackberry PPO. However, less powerful are NaF, chloride acid, citric acid, and NaSO4, which exhibited the lowest percentage of inhibition. Lineweaver-Burk plots of the blackberry PPO in the presence of ascorbic acid, citric acid, L-cysteine, acid chlorhydric, quercetin, reduced glutathione, NaF, and NaSO4 respectively are represented in (Figure 8). Furthermore, it was found that the inhibitors exhibit different types of inhibition (Figure 8).
Ascorbic acid reduces quinones to hydroquinones without direct PPO inhibition. It can prevent enzymatic browning only as long as it is present in the reduced form. This prevents the formation of key intermediates and inhibits the activity of the oxidase [40] Ascorbic acid is the best competitive inhibitor, present at 0.1 mM with a 100% inhibitory effect on the PPO of the wild blackberry. This result is similar to that of blueberry, with a 99.7% inhibition effect at 2 mM 37.
Glutathione does not appear to affect the enzyme directly, and oxygen uptake may be stimulated or inhibited depending on the particular phenol being oxidized 41. Glutathione is a potent competitive inhibitor of the wild blackberry PPO. This result is identical to that of Parsley PPO [42]. In contrast to our results, some authors reported that the inhibition type of glutathione and ascorbic acid was a mixed-type inhibition for lettuce PPO 41.
L-cysteine can readily bind to quinones to create complexes that prevent secondary oxidation and polymerization processes, which consume the substrate. It can also act as a reducing agent 43. L-Cysteine is the best non-competitive inhibitor present at 0.1 mM, with a 93.3% inhibitory effect on the blackberry PPO. This result is similar to that of blueberry, with a 99.4% inhibitory effect at 2 mM 37, and that of PPO parsley 42. Quercetin was a potent inhibitor even at a low concentration of 0.1 mM, which exhibited an 89% inhibitory effect on pyrocatechol substrate with competitive inhibition. This result is identical to that of the cape gooseberry PPO 44 with 71.7% but with a 4-methylcatechol substrate difference and uncompetitive inhibition. Sodium fluoride is a weak inhibitor of the wild blackberry PPO but is shown to be the best inhibitor of palm kernel PPO 45. Citric acid and chloride acid are weak inhibitors of the blackberry. These results are different to those of the PPO for mamey (Pouteriasapota) 46. The enzyme kinetics indicated the presence of two isoenzymes.
Table 4: Effect of inhibitors on the PPO activity and type of inhibition
| Anti-Browning agents | [I] (mM) | IC50 (mM) | Ki (mM) | Inhibition% | Type of inhibition |
| Ascorbic acid | 10 | 0.4±0.02d | 0.18± 0.00e | 100 ± 0.00a | Competitive |
| 1 | 78 ± 0.02d | ||||
| 0,1 | 29 ± 0.02fg | ||||
| Reduced glutathione | 10 | 0.2±0.00d | 0.2±0.00e | 100± 0.00a | non competitive |
| 1 | 30± 0.03fg | ||||
| 0,1 | 8 ± 0.02jk | ||||
| Citric acid | 10 | 1.2± 0.01c | 0.6±0.00d | 29 ± 0.05fg | non competitive |
| 1 | 24 ± 0.04h | ||||
| 0,1 | 17 ± 0.03i | ||||
| L-Cysteine | 10 | 1.25±0.04c | 1.25±0.06b | 93.3 ± 0.00a | non competitive mixte |
| 1 | 52 ± 0.02e | ||||
| 0,1 | 27 ± 0.02hg | ||||
| Chloride acid | 10 | 5±0.40a | 2.5±0.20a | 30 ± 0.02fg | Competitive |
| 1 | 10 ± 0.02j | ||||
| 0,1 | 5 ± 0.00k | ||||
| Quercetin | 10 | 0.38±0.02d | 0.19±0.01e | ND | Competitive |
| 1 | ND | ||||
| 0,1 | 89 ± 0.01c | ||||
| NaSo4 | 10 | 2.6±0.20b | 1.3±0.10b | 9 ± 0.00j | Competitive |
| 1 | 8.7 ± 0.01j | ||||
| 0,1 | 8.26 ± 0.00jk | ||||
| NaF | 10 | 1.2± 0.06 | 1±0.006c | 32±0.02f | Mixte |
| 1 | 11±0.04j | ||||
| 0,1 | 7.47±0.01jk |
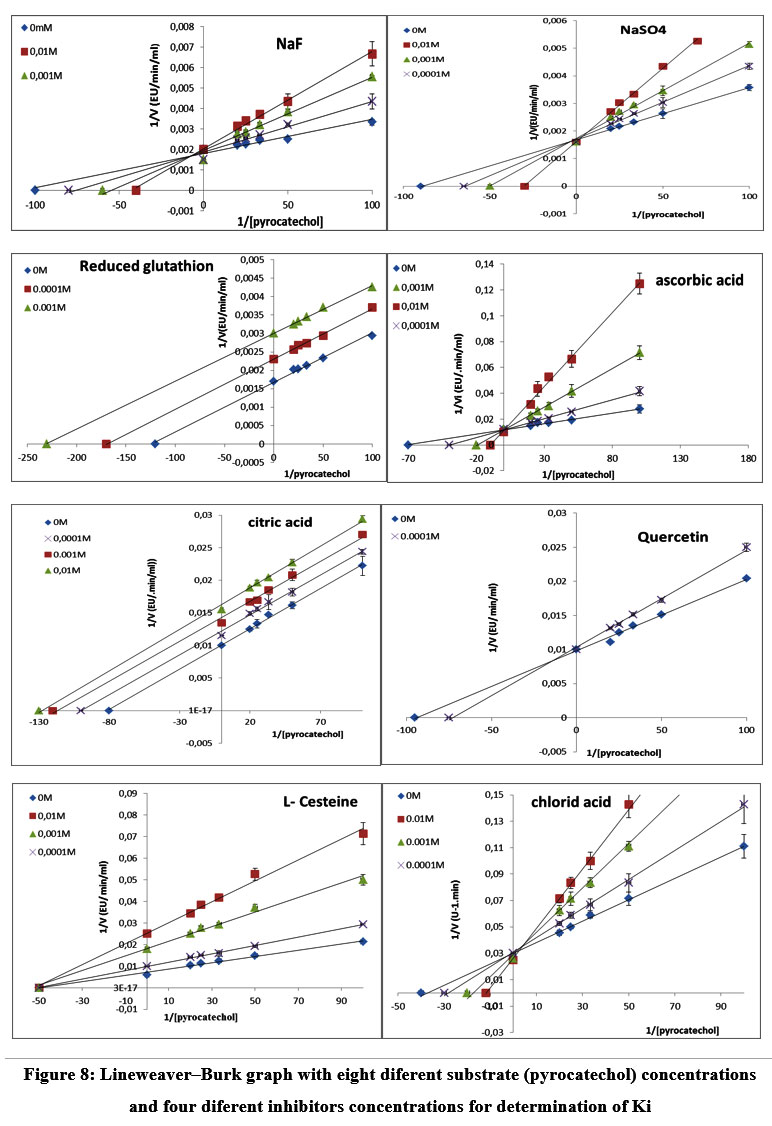 |
Figure 8: Lineweaver–Burk graph with eight diferent substrate (pyrocatechol) concentrations and four diferent inhibitors concentrations for determination of Ki. |
Effect of natural extract fresh inhibitors on enzyme activity.
Table 5: Effect of inhibitors naturals on the PPO activity and type of inhibition.
| Natural anti-Browning agents | Inhibition % | Vmax (EU/min/mL) | Km (M) | Type of inhibition |
| Onion fresh | 50%±1b | 167±0.6a | 0, 33±0.002a | Competitive |
| Arbutus 6% fresh | 60%±2a | 167±1a | 0,17±0.002b | Competitive |
The results show that the antibrowning effect of the diluted by A. Unedo fruit fresh 6% is greater than that of the onion fresh. The values in percentages of the inhibition effect show that the arbutus has a value of 60% and that the fresh onion has a value of 50%. They have the same values of Vmax with different Km values. Figure 9 (a) and 9 (b) show the Lineweaver-Burk plots of the blackberry PPO in the presence of fresh A. Unedo fruit diluted 6% and fresh onion extract, respectively. It was found that two natural agents exhibit similar types of inhibition, which were competitive and irreversible.
These two natural sources have powerful antioxidant effects; anthocyanins; phenolic compounds; and vitamin C. The anti-browning effect of A. Unedo is stronger than that of the fresh onion, which may be due to its higher vitamin C content 2. We conclude, therefore, that arbutus extract could be used as a natural food ingredient for the prevention of the browning caused by the blackberry PPO.
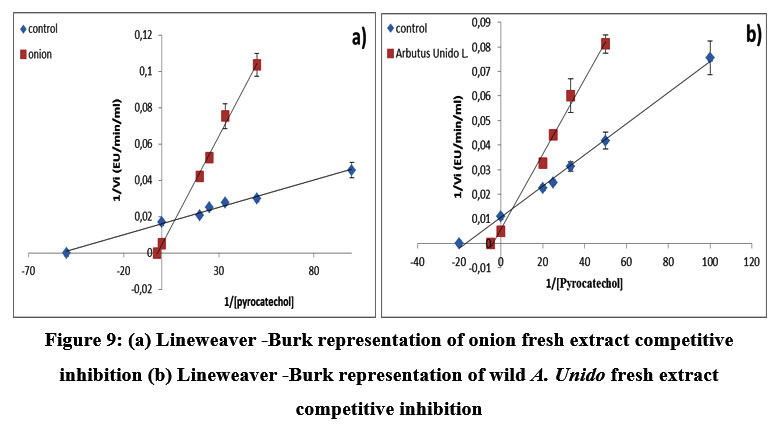 |
Figure 9: (a) Lineweaver -Burk representation of onion fresh extract competitive inhibition (b) Lineweaver -Burk representation of wild A. Unido fresh extract competitive inhibition |
Blackberry PPO showed maximum activity when observed with pyrocatechol, followed by with 4-methylcatechol. Our results were similar to blackberry 19, blueberry 37, and tea leaf (Camellia sinensis) 47. Quercetin is a stronger chemical inhibitor because it was able to inhibit 89% of our blackberry PPO at a concentration of 0.1 mM, followed by ascorbic acid, which was able to inhibit 29% at the same concentration. These two inhibitors are competitive in nature and exhibit a small ki. Onion competitively inhibits our PPO, which is similar to the yam PPO from Yapi et al. 48. Mayer claims that there are two categories of PPO inhibitors: those that bind to the copper side and those that interfere with the side of phenolic compounds. Competition-based inhibition is present in the category of interactions with copper, whereas non-competitive inhibition is present in the other category 49.
Competitive inhibition was found for the tea leaf (Camellia sinensis) with sodium sulfate, ascorbic acid, glutathione, and dithioerythritol inhibitors using catechol as a substrate 47.
Our natural extracts show a higher anti-browning effect than that provided by Lim [50], who found 47.97%. These results outperform those provided by Lim 50.
Fresh onion and wild A. Unedo extract were able to inhibit the blackberry PPO activity by up to 50% and 60%, respectively. These two natural inhibitors are competitive types therefore able to form chelate complexes with Cu in the active site of PPO, as cited by Mayer 49. These results are identical for the onion as a natural inhibitor with the same type of competitive inhibition 51. The use of chemical and natural inhibitors for controlling the blackberry PPO has not been investigated previously.
Conclusion
The polyphenol oxidase from the blackberry that leads to the enzymatic browning need to be prevented during storage and processing. PPO was purified, characterized and kinetic parameters for five substrates were pyrocatechol, 4-methylcatechol Pyrogallol, gallic acid, and tyrosine were determined. The optimum pH and temperature were pH 6.6 and 25°C, respectively. The wild blackberry PPO has two isoenzymes, one with 22 kD and 70 kD, respectively. The enzyme activity indicated variable affinity towards o-dihydroxy phenolic substrates and no activity was observed with phenol, a monohydroxy substrate, and was very effective towards pyrocatechol flowed by Pyrogallol and 4-Methyl catechol. Quercetin and Ascorbic acid have the lowest Ki values, and fresh onion and wild Arbutus extract were able to inhibit the blackberry PPO activity up to 50% and 60%, respectively. This may provide new insight into natural PPO inhibitors in the goal of replacing the chemical anti-browning agents such as sulphites with natural ones for protection human health against toxics and may contribute to increased knowledge of the natural inhibitor of PPO.
Acknowledgment
We thank Dr.Jose A. Gavira Gallardo and Carmen Lopez Sauchez for facilities at the Laboratorio de Universidad de Granada laboratory of Research Scientist of the CSIC.
Conflict of Interest
The authors declare no conflict of interest.
Funding Source
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
- Junqueira-Gonçalves, M.P., Alarcón, E., Niranjan, K. (2016).The Efficacy of Potassium Sorbate-Coated Packaging to Control Postharvest Gray Mold in Raspberries, Blackberries and Blueberries. Postharvest Biol. Technol.111, 205–208 DOI:10.1016/j.postharvbio.2015.09.014
CrossRef - Ruiz-Rodríguez, B.-M., Morales, P., Fernández-Ruiz, V., Sánchez-Mata, M.-C., Cámara M., Díez-Marqués C., Pardo-de-Santayana, M., Molina, M., Tardío, J. (2011). Valorization of wild strawberry-tree fruits (Arbutus unedo L.) through nutritional assessment and natural production data. Food Res. Int. 44, 1244–1253. Doi: 10.1016/j.foodres.2010.11.015.
CrossRef - Lee, J., & Wrolstad, R. E. (2004). Extraction of anthocyanins andpolyphenolics from blueberry processing waste. Journal of Food Science, 69 (7), 564-573. http://dx.doi.org/10.1111/j.1365-2621.2004.tb13651.x.
CrossRef - Araji S., Grammer T.A., Gertzen R., Anderson S.D., Mikulic-Petkovsek M., Veberic R., Phu M.L., Solar A., Leslie C.A., Dandekar A.M., Escobar M.A. (2014). Novel roles for the polyphenol oxidase enzyme in secondary metabolism and the regulation of cell death in walnut. Plant Physiol 164 (3):1191–1203. DOI: 10.1104/pp.113.228593
CrossRef - Yoruk, R., & Marshall, M. R. (2003). Physicochemical properties and function of plant polyphenol oxidase: A review. Journal of Food Biochemistry, 27(5), 361–422. doi:10.1111/j.1745-4514.2003.tb00289.x.
CrossRef - Mayer, A.M. (2006). Polyphenol oxidases in plants and fungi: Going places? .A review. Phytochemistry, 67 (21) 2318–2331. DOI: 10.1016/j.phytochem.2006.08.006
CrossRef - Othman, O.C. (2014). Polyphenoloxidase and perioxidase activity during open air ripening storage of pineapple (Ananas comosus L.), mango (Mangifera indica) and papaya (Carica papaya) fruits grown in Dar es Salaam, Tanzania. Tanzan J Sci 38(3):84–94. https://www.ajol.info/index.php/tjs/article/view/100182
- Yoruk, R., & Marshali, M. R. (2003). Physicochimical Propertles and Function of Plant Polyphenol oxidase a Review. Journal of Food Biochemistry, 27(5), 361–422. Doi:10.1111/j.1745-4514.2003.tb00289.x
CrossRef - Fang, Z., Zhang, M., Sun, Y., Sun, J. (2007). Polyphenol oxidase from bayberry (Myrica rubra Sieb. et Zucc.) and its role in anthocyanin degradation. Food Chem. 103, 268–273. Doi: 10.1016/j.foodchem.2006.07.044
CrossRef - Mishra, B. B., Gautam, S., & Sharma, A. (2013). Free phenolics and polyphenol oxidase (PPO): The factors affecting post-cut browning in eggplant (Solanum melongena). Food Chemistry, 139(1-4), 105–114. doi:10.1016/j.foodchem.2013.01.0
CrossRef - Constabel, C. P., Bergey, D. R., & Ryan, C. A. (1995). Systemin activates synthesis of wound-inducible tomato leaf polyphenol oxidase via the octadecanoid defense signaling pathway. Proceedings of the National Academy of Sciences, 92(2), 407–411. Doi:10.1073/pnas.92.2.407
CrossRef - Queiroz, C., Mendes Lopes, M. L., Fialho, E., and Valente-Mesquita, V. L. (2008) Polyphenol Oxidase: Characteristics and Mechanisms of Browning Control, J. Food Reviews International, 24 (4), 361–375. https://doi.org/10.1080/87559120802089332
CrossRef - Moon, K.M., Kwon, E.-B., Lee, B., and Kim, C. Y. (2020). Recent Trends in Controlling the Enzymatic Browning of Fruit and Vegetable Products, J. Molecules, 25 (12) 2754. DOI: 10.3390/molecules25122754
CrossRef - Jayawardena, E., Vanniarachchi, M., and Wansapala, J. (2019). Review on Non-thermal Technologies for the Preservation of Fruit Juices, Journal of Agricultural Science and Technology B 9, (6) 365-372. DOI:10.17265/2161-6264/2019.06.001
CrossRef - Rawson, A., Patras, A., Tiwari, B. K., Noci, F. Koutchma, T., and Brunton, N. (2011) Effect of thermal and non thermal processing technologies on the bioactive content of exotic fruits and their products: Review of recent advances, J. Food Research International 44(7) 1875–1887. https://doi.org/10.1016/j.foodres.2011.02.053
CrossRef - Oms-Oliu, G., M. A. Rojas-Graü, L. A. González, P. Varela, R. Soliva-Fortuny, Hernando, M. I. H.,… and Martín-Belloso, O. (2010). Recent approaches using chemical treatments to preserve quality of fresh-cut fruit: A review. Postharvest Biology and Technology, 57(3), 139–148. Doi:10.1016/j.postharvbio.2010.04.001
CrossRef - Carocho, M., M. F. Barreiro, P. Morales, and I. C. F. R. Ferreira. (2014). Adding Molecules to Food, Pros and Cons: A Review on Synthetic and Natural Food Additives. Comprehensive Reviews in Food Science and Food Safety, 13(4), 377–399. Doi:10.1111/1541-4337.12065.
CrossRef - Loizzo, M. R., R. Tundis, M. Bonesi, F.Menichini, D. De Luca, C. Colica, and F. Menichini. (2012). Evaluation of Citrus aurantifoliapeel and leaves extracts for their chemical composition, antioxidant and anti-cholinesterase activities. Journal of the Science of Food and Agriculture, 92(15), 2960–2967. Doi:10.1002/jsfa.5708.
CrossRef - González, E. M., B. de Ancos, and M. P. Cano. (2000). Partial Characterization of Peroxidase and Polyphenol Oxidase Activities in Blackberry Fruits. Journal of Agricultural and Food Chemistry, 48(11), 5459–5464. Doi: 10.1021/jf000169w.
CrossRef - LEE, M.-K., KIM, Y.-M., KIM, N.-Y., KIM, G.-N., KIM, S.-H., BANG, K.-S., & PARK, I. (2002). Prevention of Browning in Potato with a Heat-treated Onion Extract. Bioscience, Biotechnology, and Biochemistry, 66(4), 856 858.Doi:10.1271/bbb.66.856 10.1271/bbb.66.856
CrossRef - Kim, M.-J., Kim, C. Y. and I. (2005). Park, Prevention of enzymatic browning of pear by onion extract, Food Chemistry, 89 (2), 181–184. DOI: 10.1016/j.foodchem.2004.02.018
CrossRef - Lee, J., and Wrolstad, R. E. (2006). Extraction of Anthocyanins and Polyphenolics from Blueberry Processing Waste. Journal of Food Science, 69(7), 564–573. Doi:10.1111/j.1365-2621.2004.tb13651.x
CrossRef - Yaǧar, H., and A. Saǧiroǧlu. (2002). Partially purification and characterization of polyphenol oxidase of quince, Turkish Journal of Chemistry. 26(1) 97-103. https://journals.tubitak.gov.tr/chem/vol26/iss1/11.
- Davis, B. J. (1964). Disc electrophoresis II. Method and application to human serum proteins, Annals of the New York Academy of Science, 121, 404-27. Doi: 10.1111/j.1749-6632.1964.tb14213.x
CrossRef - Bradford, M. M., (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72(1-2), 248–254. Doi:10.1016/0003-2697(76)90527-3.
CrossRef - Lineweaver, H., and Burk, D. (1934). The Determination of Enzyme Dissociation Constants. Journal of the American Chemical Society, 56(3), 658–666. Doi: 10.1021/ja01318a036.
- Wong, C. W. and P. L. Angel Lee. (2014). Inhibitory effect of onion extract on cassava leaf (Manihot esculenta Crantz) polyphenol oxidase, International Food Research Journal. 21(2) 755-758.
CrossRef - Yang, C.P., Fujita, S., Ashrafuzzaman, M., Nakamura, N., and Hayashi, N. (2000). Purification and characterization of polyphenol oxidase from banana (Musa sapientum L.) pulp, J. Agric. Food Chem. 48 (7) 2732–2735. Doi: 10.1021/jf991037+.
CrossRef - Kumar, V. B. A., Mohan, T. C. K., and Murugan, K. (2008). Purification and kinetic characterization of polyphenol oxidase from Barbados cherry (Malpighia glabra L.). Food Chemistry, 110(2), 328–333. Doi:10.1016/j.foodchem.2008.02.006.
CrossRef - Ding, C.-K., Chachin, K., Ueda, Y., and Imahori, Y. (1998). Purification and Properties of Polyphenol Oxidase from Loquat Fruit. Journal of Agricultural and Food Chemistry, 46(10), 4144–4149. Doi: 10.1021/jf980093s.
CrossRef - Ziyan, E., and Perkyardimci, S. (2004). Purification and characterization of Pear (Pyrus communis) Polyphenol Oxidase, Turkish Journal of Chemistry. 28 547–557. Doi: 10.1134/S0033994619020134.
CrossRef - Nagai, T., and Suzuki, N. (2001). Partial Purification of Polyphenol Oxidase from Chinese Cabbage Brassica Rapa L. Journal of Agricultural and Food Chemistry, 49(8), 3922–3926. Doi: 10.1021/jf000694v.
CrossRef - Gawlikdziki, U., Szymanowska, U., and Baraniak, B. (2007). Characterization of polyphenol oxidase from broccoli (Brassica oleracea var. botrytis italica) florets. Food Chemistry, 105(3), 1047–1053. Doi:10.1016/j.foodchem.2007.05.012.
CrossRef - Bibhuti, B. Mishra, Satyendra Gautam, Arun Sharma (2012). Purification and characterisation of polyphenol oxidase (PPO) from eggplant (Solanum melongena). Food Chemistry, 134(4), 0–0. doi:10.1016/j.foodchem.2012.03.098
CrossRef - Wititsuwannakul, D. (2002). Polyphenol oxidases from latex of Hevea brasiliensis: purification and characterization. Phytochemistry, 61(2), 115–121. Doi:10.1016/s0031-9422(02)00234-0.
CrossRef - Liu, N., Liu, W., Wang, D., Zhou, Y., Lin, X., Wang, X., and Li, S. (2013). Purification and partial characterization of polyphenol oxidase from the flower buds of Lonicera japonica-Thunb. Food Chemistry, 138(1), 478–483. Doi:10.1016/j.foodchem.2012.10.103.
CrossRef - Siddiq, M., and Dolan, K. D. (2017). Characterization of polyphenol oxidase from blueberry (Vaccinium corymbosum L.). Food Chemistry, 218, 216–220. Doi:10.1016/j.foodchem.2016.09.061
CrossRef - Montero, P., Ávalos, A., and Pérez-Mateos, M. (2001). Characterization of polyphenoloxidase of prawns (Penaeus japonicus). Alternatives to inhibition. Food Chemistry, 75(3), 317–324. Doi:10.1016/s0308-8146(01)00206-0.
CrossRef - Kavrayan, D., and Aydemir, T. (2001). Partial purification and characterization of polyphenoloxidase from peppermint (Mentha piperita). Food Chemistry, 74(2), 147–154. Doi: 10.1016/s0308-8146(01)00106-6.
CrossRef - Schwartz, B., Olgin, A. K., and Klinman., J. P. (2001). The Role of Copper in Topa Quinone Biogenesis and Catalysis, as Probed by Azide Inhibition of a Copper Amine Oxidase from Yeast. Biochemistry, 40(9), 2954–2963. Doi:10.1021/bi0021378.
CrossRef - Doğru, Y. Z., and Erat, M. (2012). Investigation of some kinetic properties of polyphenol oxidase from parsley (Petroselinum crispum, Apiaceae). Food Research International, 49(1), 411–415. Doi:10.1016/j.foodres.2012.07.028.
CrossRef - Doğru, Y. Z. and Erat, M. (2012). Investigation of some kinetic properties of polyphenol oxidase from parsley (Petroselinum crispum, Apiaceae), J. Food Research International, 49(1) (2012) 411–415. Doi : 10.1016/j.foodres.2012.07.028
CrossRef - Wesce-Ebeling, P., and Montgomery, M. W. (1990). Strawberry Polyphenoloxidase: Extraction and Partial Characterization. Journal of Food Science, 55 (5), 1320–1324. Doi:10.1111/j.1365-2621.1990.tb03925.x.
CrossRef - Bravo, K., and E. Osorio. (2016). Characterization of polyphenol oxidase from Cape gooseberry (Physalis peruviana L.) fruit. Food Chemistry, 197, 185–190. Doi:10.1016/j.foodchem.2015.10.126.
CrossRef - Robert, C., C. Rouch, and F. Cadet. (1997). Inhibition of palmito (Acanthophoenix rubra) polyphenol oxidase by carboxylic acids. Food Chemistry, 59 (3), 355–360. Doi:10.1016/s0308-8146(96)00179-3.
CrossRef - Palma-Orozco, G., Sampedro, J. G. A., Ortiz-Moreno, and Nájera , H. (2012). In situ Inactivation of Polyphenol Oxidase in Mamey Fruit (Pouteria sapota) by Microwave Treatment, Journal of Food Science, 77 (4) C359–C365. Doi: 10.1111/j.1750-3841.2012.02632.x
CrossRef - Öztürk, C., Aksoy, M., Küfrevioğlu, Ö. İ. (2019). Purifcation of tea leaf (Camellia sinensis) polyphenol oxidase by using afnity chromatography and investigation of its kinetic properties, Journal of Food Measurement and Characterization, 14, 31–38. Doi:10.1007/s11694-019-00264-8
CrossRef - Yapi, J.C., Gnangui, S.N., Dabonne, ´ S. (2015). Inhibitory effect of onions and garlic extract on the enzymatic browning of an edible yam (Dioscorea cayenensis-rotundata cv. Kponan) cultivated in Côte d Ivoire. Int J Curr Res Acad Rev 3(1):219–231.https://www.semanticscholar.org/paper/Inhibitory-Effect-of-Onions-and-Garlic-Extract-on-Yapi-Gnangui/1133fa09da52c97eea3c4efea1953b5a084ace6f
CrossRef - Mayer, A.M., Harel, E. (1979). Polyphenol Oxidases in Plants. Phytochemistry 18 (2) 193–215. 49. https://doi.org/10.1016/0031-9422(79)80057-6.
CrossRef - Lim, W. Y., & Wong, C. W. (2018). Inhibitory effect of chemical and natural anti-browning agents on polyphenol oxidase from ginger (Zingiber officinale Roscoe). Journal of Food Science and Technology, 55(8), 3001–3007. doi:10.1007/s13197-018-3218-7
CrossRef - Yuniarti, T., Sukarno, Yuliana, N.D., Budijanto, S. (2018). Inhibition of enzymatic browning by onion (Allium cepa L.): investigation on inhibitory mechanism and identification of active compounds, Current Research in Nutrition and Food Science, 6 (3) , 770-780. Doi:10.12944/CRNFSJ.6.3.19
CrossRef
Accepted on: 19 Sep 2022
Second Review by: Nige Show , Sofia Agriopoulou Greece
Final Approval by: Dr Prasad Shekhar Variyar





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








