Carotenoids: from Plants to Food Industry
Technological Educational Institute of Peloponnese Dept. Food Technology
Corresponding Author Email: theovarzakas@yahoo.gr
DOI : http://dx.doi.org/10.12944/CRNFSJ.4.Special-Issue1.04
Download this article as:
![]()
Carotenoids have been studied for their ability to prevent chronic disease due to the free radical theory of aging in chronic disease etiology. β-carotene, lycopene, zeaxanthine and others carotenoids have antioxidant properties, but the antioxidant capability is variable depending on the in vitro system used The physiology, structure and biochemistry is well described.
Moreover, sources of carotenoids and health effects along with bioavailability-absorption and metabolism, of carotenoids are well addressed. The effect of carotenoids on biotechnology and the food industry is significantly attributed.
Finally, carotenoids as fortified substances in foods and special aspects about carotenenoids as health promoters are well presented along with a glance of carotenoids economics.
Carotenoids; Phtochemicals; Physiology; Biochemistry; Structure; Health effects; Biotechnology
Introduction
The Carotenoids belong to phytochemicals
Phytochemicals are a large group of plant-derived compounds hypothesized to be responsible for much of the disease protection conferred from diets high in fruits, vegetables, beans, cereals, and plant-based beverages such as tea and wine. Phytochemicals and antioxidant constituents in plant material have raised interest among scientists, food manufacturers, producers, and consumers for their roles in the maintenance of human health. Dietary modification by increasing the consumption of a wide variety of fruits, vegetables, and whole grains daily is a practical strategy for consumers to optimize their health and to reduce the risk of chronic diseases.1 Phytochemical extracts from fruits and vegetables have strong antioxidant and antiproliferative activities, and the major part of total antioxidant activity is from the combination of phytochemicals. Among of phytochemicals the Carotenoids have an important position. Carotenoids are nature’s most widespread pigments and have also received substantial attention because of both their provitamin and antioxidant roles. They are used extensively as safe, natural colorants for food, feed, and cosmetics. They are known to be essential for plant growth and photosynthesis, and are a main dietary source of vitamin A in humans.2 Because humans are unable to synthesize vitamin A de novo from endogenous isoprenoids precursors, plant carotenoids provide the primary dietary source of provitamin A. Carotenoids owe their name to carrots (Daucus carota), and xanthophylls (originally phylloxanthins) are derived from the Greek words for yellow (xanthos) and leaf (phyllon). Together with anthocyanins, carotenoids are the most complex class of natural food colorants with over 750 different structures identified.
Physiology, Structure and Biochemistry
Plant chloroplasts have a remarkably similar carotenoid composition in photosynthetic tissues, with lutein (45% of the total), b-carotene (25–30%), violaxanthin (10–15%), and neoxanthin (10–15%) as the most abundant carotenoids.3 Most carotenoids are located, together with chlorophylls, in functional pigment-binding protein structures embedded in photosynthetic (thylakoid) membranes. Thus several compounds of carotenoids are involved in photosynthesis Carotenoids assist in photosynthesis by capturing energy from light of wavelengths that are not efficiently absorbed by chlorophylls. A typical carotenoid is β-carotene, whose two carbon rings are connected by a chain of 18 carbon atoms with alternating single and double bonds. Splitting a molecule of β-carotene into equal halves produces two molecules of vitamin A. Oxidation of vitamin A produces retinal, the pigment used in vertebrate vision. This explains why carrots, which are rich in β-carotene, enhance vision.4 Xanthophylls are accessory pigments in the light-harvesting antennae of the chloroplasts, which are capable of transferring energy to the chlorophylls. They also quench triplet excited states in chlorophyll molecules by dissipating the excess excitation energy in a non-radiative manner, a process known as nonphotochemical quenching (NPQ) (reviewed in.5,6 This function is crucial to protect against chlorophyll bleaching in intense light. An additional important role of carotenoids in plants is to furnish flowers and fruits with distinct colors that are designed to attract animals. In chloroplasts, carotenoids play vital roles in photosynthesis and are indispensable, whereas in chromoplasts, they can be considered as secondary metabolites. Carotenoids in plants are also precursors for the synthesis of the hormone abscisic acid (ABA).7,8
The carotenoids as above mentioned are isoprenoid compounds, biosynthesized by tail-to-tail linkage of two C20 as seen in (Fig. 1), forming a complex of 40-carbon skeleton of isoprene units (Fig. 2). The most striking and characteristic feature of carotenoids structure as that is illustrated in Fig 1&2 is the long stem of alternate single and double bonds that forms the central part of the molecule. This constitutes a conjugated system in which the п-electrons are effectively delocalized over the entire length of the polyene chain. It is the feature that gives the carotenoids as a group their distinctive molecular shape, chemical reactivity and light absorbing properties. Carotenoids are lipophilic, but some of them with polar hydroxyl and keto functionalities, as seen in fig 2, have increased affinities for lipid/water interfaces. Thus, the binding and structural traits with polar hydroxyl and keto– units of the molecules in relation to the water/lipid interface is considered important for the antioxidant efficiency of carotenoids.
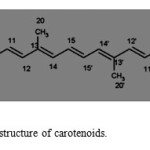 |
Figure 1: Basic chemical structure of carotenoids. Click here to View figure |
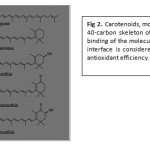 |
Figure 2: Carotenoids, molecular structures of 40-carbon skeleton of isoprene units. The binding of the molecules at the water/lipid interface is considered important for the antioxidant efficiency. |
The structure may be cyclized at one or both ends, may have various hydrogenation levels, or may possess oxygen-containing functional groups. Lycopene and carotene are examples of acyclized and cyclized carotenoids, respectively. Carotenoids compounds most commonly occur in nature in the all-trans form. The most characteristic feature of carotenoids is the long series of conjugated double bonds forming the central part of the molecule. This gives them their shape, chemical reactivity, and light-absorbing properties. Carotene, β-carotene, and-cryptoxanthin are able to function as provitamin A. Each enzymatic step from phytoene to lycopene adds one double bound to the molecule, resulting in lycopene, which is a symmetrical molecule containing 13 double bonds. The biosynthetic step after lycopene involves enzymatic cyclization of the end groups, which results in γ-carotene (one β- ring) and β-carotene (two β– rings). The addition of oxygen to the molecule leads to the formation of xanthophylls.3 The concentration of each carotenoids in a fruit or vegetable suggests which enzyme or enzymes may be rate-limiting in the biosynthetic cascade. For example a very high concentration of lycopene in red tomatoes suggests a lack of sufficient enzyme activity to convert lycopene to γ-carotene (i.e. insufficient cyclase activity).9 The biosynthetic sequence of the carotenoids in plants seems simple in fig 3 and with details in Fig 4, by Botella-Pavia and Rodriguez-Concepcion.10
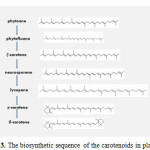 |
Figure 3: The biosynthetic sequence of the carotenoids in plants. |
 |
Figure 4: Carotenoids biosynthesis and related pathways in plants. Click here to View figure |
Botella-Pavia P and RodriguezConcepcion M (2006) Carotenoid biotechnology in plants for nutritionally improved foods.Physiol Plant 126:369–381 GAP, glyceraldehyde 3-phosphate; DXP, deoxyxylulose 5-phosphate; MEP, methylerythritol 4-phosphate; CDP-ME, 4-diphosphocytidyl-methylerythritol; CDP-MEP, 4-diphosphocytidylmethylerythritol 2-phosphate; ME-cPP, methylerythritol 2,4-cyclodiphosphate; HMBPP, hydroxymethylbutenyl 4-diphosphate; IPP, isopentenyl diphosphate; DMAPP, dimethyl allyl diphosphate; GPP, geranyl diphosphate; GGPP, geranylgeranyl diphosphate; ABA, abscisic acid. Enzymes are indicated in bold: DXS, DXP synthase; DXR, DXP reductoisomerase; CMS, CDP-ME synthase; CMK, CDP-ME kinase; MCS, ME-cPP synthase; HDS, HMBPP synthase; HDR, HMBPP reductase; IDI, IPP isomerase; GGDS, GGPP synthase; PSY, phytoene synthase; PDS, phytoene desaturase; ZDS, z-carotene desaturase; CRTISO, carotenoid isomerase; LCYB, lycopene b-cyclase; LCYE, lycopene e-cyclase; CHYB, carotenoid b-ring hydroxylase; CHYE, carotenoid e-ring hydroxylase; ZEP, zeaxanthin epoxidase; VDE, violaxanthin de-epoxidase; NSY, neoxanthin synthase.
The deficiency of vitamin A, which remains one of the most noticeable nutritional problems in many parts of the world, affects an estimated 250 million preschool children and causes blindness in up to 500,000 of them annually.11 Apart from the nutritional significance, carotenoids as antioxidants have been implicated in reducing the risk of cancer and cardiovascular diseases.12 Some carotenoids also offer protection against age related eye diseases, such as macular degeneration, the leading cause of blindness in the elderly.13 Thus, development of carotenoids enriched food crops provides the most effective and sustainable approach to maximize the nutritional and health benefits of carotenoids to a large number of population in the world.
Sources of carotenoids and health effects
Apricots, cantaloupe, carrots, pumpkin and sweet potato are sources of a-carotene and b-carotene; pink grapefruit, tomatoes and watermelon are sources of lycopene, z-carotene, b-carotene, phytofluene and phytoene. Mango, papaya, peaches, prunes, squash and oranges are sources of lutein, zeaxanthin, and b-cryptoxanthin, a-, b- and z-carotene, phytofluene and phytoene, whereas green fruits and vegetables such as green beans, broccoli, brussel sprouts, cabbage, kale, kiwi, lettuce, peas and spinach are sources of lutein, zeaxanthin, a- and b-carotene. Common dietary sources of carotenoids in regular vegetable foods (μg/100 fresh weight) are illustrated in Table 1. Carotenoid concentrations in fruits and vegetables vary with plant variety, degree of ripeness, time of harvest, and growing and storage conditions.14 The most widely studied carotenoids related to health effects are β-carotene, lycopene, lutein, zeaxanthin, β-cryptoxanthin, and α-carotene. β-carotene is associated with reduction of cardiovascular disease risk,15,16 while α-carotene is recognized as an anticarcinogenic agent.17,18 Lycopene is the carotenoid with greater biological efficiency to scavenge singlet oxygen19 and may have protective effect against some types of cancer.20,21,22 Lutein and zeaxanthin are the yellow macular pigments and act as antioxidants23 and blue light filters of high energy.24 β-cryptoxanthin has been associated with antimutagenic activities in in vitro studies,25 with immunomodulary activities on in vitro and in vivo studies,26 and with effects on osteosynthesis in in vitro and in vivo studies.27
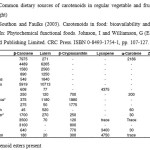 |
Table 1: Common dietary sources of carotenoids in regular vegetable and fruits (μg/100 fresh weight) Source: Southon and Faulks (2003). |
Carotenoids in food: bioavailability and functional benefits. In: Phytochemical functional foods. Johnson, I and Williamson, G (Eds.). Ch. 7. Woodhead Publishing Limited. CRC Press. ISBN 0-8493-1754-1, pp. 107-127.
Bioavailability – Absorption and Metabolism, of Carotenoids
The bioavailability of carotenoids in foods and in commercial preparations varies widely.87 Only about 5% of the carotenoids in whole, raw vegetables, for example, is absorbed by the intestine, whereas 50% or more of the carotenoids is absorbed from micellar solutions. Thus, the physical form in which the carotenoids is presented to intestinal mucosal cells is of crucial importance.27
Food matrix
Carotenoids have an essential physiological function as a vitamin A precursor. The release of carotenoids from food matrix, their dispersion within the digestive tract, and their solubilization in mixed micelles are important steps for carotenoids bioaccessibility. Solubilized carotenoids are taken up by epithelial cells of the small intestine by simple diffusion and/or transporter-mediated processes and then secreted to lymph as chylomicron. Carotenoids are incorporated into triglyceride-rich lipoproteins and released into the circulation.28 Triglycerides are depleted from circulating chylomicrons through the activity of an enzyme called lipoprotein lipase, resulting in the formation of chylomicron remnants. Chylomicron remnants are taken up by the liver, where carotenoids are incorporated into lipoproteins and secreted back into the circulation. This is carried out in the presence of fat and conjugated bile acids. Chylomicrons are responsible for the transport of carotenoids from the intestinal mucosa to the bloodstream via the lymphatics for delivery to tissues. Carotenoids are transported in the plasma exclusively by lipoproteins. Absorption is affected by the same factors that influence fat absorption. Thus, the absence of bile or any generalized malfunction of the lipid absorption system, such as diseases of the small intestine and pancreas, will interfere with the absorption of carotenoids. Carotenoids accumulated in tissues are thought to be metabolized to small molecules by enzymatic cleavage and/or chemical oxidation with active oxygen species at conjugated double bonds. The hydroxyl group of xanthophylls can be oxidatively metabolized to carbonyl group. Carotenoids with long chain of conjugated double bonds physically quench singlet oxygen and scavenge oxygen radicals, particularly under low oxygen pressure, and thereby they have been thought to work as lipophilic antioxidants for human health. Carotenoids, being mostly fat soluble, follow the same intestinal absorption path as dietary fat. For carotenoid absorption, as little as 3-5 g of fat in a meal is sufficient.29,30 Oxygen functionalized carotenoids are more polar than carotenes. Thus, α-carotene, β-carotene and lycopene tend to predominate in low-density lipoproteins (LDL) in the circulation, whereas high-density lipoproteins (HDL) are major transporters of xanthophylls such as cryptoxanthins, lutein and zeaxanthin.31,32 The delivery of carotenoids to extrahepatic tissues is accomplished through the interaction of lipoprotein particles with receptors and the degradation by lipoprotein lipase. Although no less than forty carotenoids are usually ingested in the diet, only six carotenoids and their metabolites have been found in human tissues, suggesting selectivity in the intestinal absorption of carotenoids.33,34 In contrast, thirty-four carotenoids and eight metabolites are detected in breast milk and serum of lactating mothers.35 Recently, facilitated diffusion in addition to simple diffusion has been reported to mediate the intestinal absorption of carotenoids in mammals. The selective absorption of carotenoids may be due to uptake to the intestinal epithelia by means of facilitated diffusion and an unknown mechanism of excretion into the intestinal lumen. It is well known that β-carotene can be metabolized to vitamin A after intestinal absorption of carotenoids, but little is known about the metabolic transformation of non-provitamin A xanthophylls. The enzymatic oxidation of the secondary hydroxyl group leading to keto-carotenoids would occur as a common pathway of xanthophyll metabolism in mammals.33 However there are some cases where the bioavailability dependent on plant sources or from fiber of meals:
- The bioavailability of carotenoids from various plant sources is thought to differ (although the degree is uncertain) due to differences in plant matrices and content of inhibitors of carotenoid absorption. For example, de Pee,36 who studied anemic school children, observed that among β-carotene-rich foods, orange-pigmented fruit was superior to dark-green, leafy vegetables in increasing circulating concentrations of β-carotene. Ripe fruits, in general, have chromoplasts that store carotenoids, and carotenoids would be expected to be more easily liberated from chromoplasts than from chloroplasts of dark-green, leafy vegetables. Of course, many of the foods that consumers think of as vegetables [e.g., tomatoes, peppers (Capsicum annuum L.)] are, botanically, fruits.
- Dietary fiber can reduce the bioavailability of carotenoids, as demonstrated with subjects who ate small meals that were either low in fiber or contained 12 g of citrus pectin.37 Immediately following the meal, the subjects ingested 25 mg of β- carotene as a supplement. Thirty hours after the treatment, the subjects’ plasma β-carotene concentrations increased 141% and 60%, respectively, for the low- and high-fiber treatments.
Interactions among carotenoids
More, there are studies about interactions among carotenoids. It is generally assumed that large supplemental doses of a single carotenoid will affect the bioavailability of other carotenoids. One carotenoid may enhance the absorption of another, compete with another for absorption, spare another, or otherwise alter the rate of metabolism of other carotenoids. For example, absorption (plasma response) of canthaxanthin38 and lutein39,40 seems to be inhibited by concurrent ingestion of b-carotene. The impact of lutein on b-carotene is less clear; lutein reduced the plasma response of b-carotene for some subjects, but increased it for others.41 In another study, lutein, but not lycopene, inhibited b-carotene absorption when administered with an equivalent amount (15 mg) of β-carotene42 Higher plasma a-carotene concentrations were observed in subjects receiving b-carotene supplements (30 mg daily for 6 weeks).40 Combined ingestion of β-carotene and lycopene resulted in greater lycopene absorption, as indicated by plasma response, than did a single dose of lycopene.41 These data indicate that b-carotene, a hydrocarbon carotene, may inhibit the absorption of oxycarotenoids, such as lutein and canthaxanthin, but enhances absorption of hydrocarbon carotenes, such as a-carotene and lycopene.
Influence of isomeric form on bioavailability and bioactivity
Among the factors that influence bioavailability and bioactivity are the food matrix, the growth maturity when the fruits and vegetable are consumed fresh and the food processing methods. The concentration of carotenoids isomers consists another serious factor because the carotenoids, exist as trans and cis-isomers.
In food research, carotenoids from fruits and vegetables have attracted a great deal of attention, mainly focusing on the analysis of geometric carotenoid isomers. There is some evidence that isomeric form may be an important factor in the bioactivity of certain carotenoids. For instance, cis-lycopene is found at high levels in human serum and tissues, while in most plant tissues and foods, the all-trans form predominates unlike lycopene,42 the trans isomer of lutein is the predominant form found both in raw plant materials and some human tissues, and has also been found to be preferentially absorbed by cell cultures.43 According to some other findings the cis-lycopene isomers concentrations are higher in animal tissues than in raw plant materials, has created among the nutritionists a dialogue as to whether cis isomers may be preferentially absorbed by the body or trans lycopene is converted to cis once consumed .44,45,46,47,48,49 Bile acid micelles mixed with lycopene isomers were found to have higher incorporation levels of cis isomers.46 In,48 high plasma lycopene content was determined using human subjects fed a sauce made of tangerine tomatoes. However, this feeding had high levels (97%) of cis isomers so it was added to the support of the hypothesis that cis lycopene isomers are more bioavailable than all-trans isomers. Another study by cis isomers of beta-carotene has also been reported to have greater efficiency of incorporation into micelles than all-trans beta-carotene in vitro50,51 but were not found to be preferentially absorbed by Caco-2 human intestinal cells.51 Difference in carotenoids solubility and shape may help explain their differences in bioavailability.49
The bioavailability of carotenoids is affected by a number of factors, heat treatment being one of them that may enhance carotenoid bioavailability.30 The reasoning behind this phenomenon is that the heating increases surface area, and agitation typically associated with thermal processing is likely to cause the breakdown of the cellular matrix of the plant material and may also induce trans to cis isomerization.44,52,53,54,55,56,58 Naturally, most of the carotenoids occur as trans-isomer in plants. However, cis-isomers may increase due to the isomerization of the trans-isomer of carotenoids during food processing.57 Relative studies8 with carrot juice processing have been carried out aiming to study the phenomenon of isomerization of carotenoids. Carrot juice is a vegetable juice and represents a rich source of natural β- and a-carotene and more. Its production is a model of processing because involves various technological steps such as blanching, acidification, pasteurization or canning. According to the studies it seems that in processing of carrot juice a partial conversion of the all-trans-carotenes towards their cis-isomers is caused.
Carotenoids and biotechnology
Carotenoids are highly beneficial for human nutrition and health because they provide
essential nutrients and important antioxidants in our diets. However, many food crops, especially the major staple crops contain only trace to low amounts of carotenoids. Although significant progress has been made in developing food crops rich in carotenoids by altering the expression of carotenoids biosynthetic genes, in many cases it has proved to be difficult to reach the desired levels of carotenoids enrichment.10 Manipulation of carotenoids content in food crops has been primarily focused on engineering of catalytic activity of this pathway by altering the expression of carotenoids biosynthetic genes. In some cases, this is not sufficient to enhance carotenoids contents to levels required for optimal human nutrition and health. A alternative approach to enhance carotenoids levels in food crops is based on regulating the formation of metabolic sinks to effectively sequester carotenoids.60 Such sinks exert their positive effect on carotenoids accumulation by pulling the pathway toward completion. This approach, in combination with manipulation of catalytic activity of the pathway, may prove to be an effective and efficient strategy to dramatically enrich carotenoids content in low-pigmented tissues of food crops. The recent identification and characterization of a novel gene mutation in cauliflower reveals that creating a metabolic sink to sequester carotenoids is an important mechanism to control carotenoids accumulation in plants. The successful demonstration of increased carotenoids accumulation in association with the formation of sink structures in transgenic crops offers a new and alternative approach to increase carotenoids content. Manipulation of the formation of metabolic sink along with the catalytic activity of the pathway may represent a promising strategy for maximally improving the nutritional quality of food crops.60 Another side of topic “biotechnology and carotenoids” is this of biotechnology as “tool” that gets involved in carotene production by manipulation of microorganisms. Natural β-carotene is widely used as a food colorant. The global market of β-carotene has been estimated to surpass USD $280 million in 2015 .61 β-carotene is very attractive as natural food colorant due to its antioxidant and pro-vitamin activities which provide additional value to the products.62 Furthermore, β-carotene was proved to have beneficial influence on iron and zinc bioaccessibilities.63 The main sources of natural β-carotene include extraction from vegetable resources and microbial fermentation.61 The production of natural colorants through fermentation has a number of advantages, such as cheaper production, higher yields, possibly easier extraction, less batch-to-batch variations, no lack of raw materials, and no seasonal variations.64,65 β-carotene can be produced by numerous microorganisms such as Blakeslea trispora (fungi), Rhodotorula spp., and Saccharomyces cerevisiae (yeast), and Dunaliella bardawil (microalgae).66,67 Although there are also some bacteria species which produce β-carotene as main carotenoid,68 these species must have the central metabolism inhibited by inorganic salts and urea or must be genetically engineered.61
Carotenoids in Food industry
Carotenoids as fortified substances in foods
In general, carotenoids are not strong antioxidants when added to foods.69 Despite all these references an additional challenge to using carotenoids as ingredients in functional foods is their high melting point, making them crystalline at food storage and at the body temperatures. Endogenous carotenoids in foods are generally stable. However, as food additives, carotenoids are relatively unstable in food systems because they are susceptible to light, oxygen,and autooxidation.57 In addition, dispersion of carotenoids into ingredient systems can result in their rapid degradation.70,71 Carotenoids can be degraded by reactions that cause the loss of double bonds or the scission of the molecule. In addition, the double bonds in carotenoids can undergo isomerization to the cis configuration.57 Isomerization reactions might actually be beneficial since cis isomers of carotenoids such as lycopene are thought to be more bioavailable and bioactive .58 Carotenoids as natural pigments, are used by the industry as colorants in various food and drinks. In literature carotenoids have been reported to act as chain breaking antioxidants under specific conditions. However due to their highly conjugated structure, carotenoids are very unstable and can be easily degraded when exposed to oxygen or light during storage or manufacture of foods. This can cause the loss of their nutritive and biological desirable properties as well as the production of undesirable flavor or aroma compounds. For these reasons, these compounds are not usually handled in their crystalline form but rather as encapsulated forms. Encapsulation of carotenoids (β-carotene) is carried out which has created an opportunity for the development of cartenoid (β-carotene) forms for supplementation and food fortification. Simple oil-in-water emulsions are currently the most widely used method of encapsulating lipophilic functional components, such as flavors and bioactive lipids. The potential for using multiple emulsions to encapsulate some lipophilic functional food or nutraceutical compounds has been demonstrated, such as β-carotene.72 The case of β-carotene b is an important member of the carotenoids family as a retinol precursor with a high conversion rate, provides a substantial proportion of vitamin A in the human diet.73,74 For these reasons, there is a strong interest in using b-carotene and other carotenoids as functional ingredients in food products. However, b-carotene is insoluble in water and weakly soluble in oil at ambient temperature because of its crystalline form, thus making it difficult to incorporate in food products and with less bioavailability.75 Furthermore, b-carotene is sensitive to light, oxygen, and heat, which limit even more its applications in the food industry.76 Recently, nanotechnology quickly emerged as one of the most promising and attractive research fields, with applications ranging from the aerospace to health industries.78 Thus the Food Industry could be have benefits from nanotechnology applications because this technology offers the potential to improve bioavailability and solubility of different functional ingredients such as the “carotenoids”.79
Special aspects about carotenenoids as health promoters
Carotenoids are thought to have a protective effect against degenerative conditions such as cancer, cardiovascular disease and cataracts .80,81 The mechanism(s) by which carotenoids exert their health benefits are not completely understood, but may be due in part to their antioxidant activities81,82 it seems that their action is supported by the common chemical feature of carotenoids where there isn’t else from the polyene chain. Polyene chain a long conjugated double bond system forming the backbone of the molecule. This chain may be terminated by cyclic end groups that contain oxygen-bearing substitutes. Many epidemiologic studies have associated high carotenoid intake with a decrease in the incidence of chronic disease. Multiple possibilities exist: certain carotenoids 1) can be converted to retinoids (i.e. have provitamin A activity), 2) can modulate the enzymatic activities of lipoxygenases (proinflammatory and immunomodulatory molecules), 3) can have antioxidants properties which are well above what is seen with vitamin A, 4) can activate the expression of genes which encode the message for production of a protein, connexin 43, which is an integral component of the gap junctions required for cell to cell communication.83 Such gene activation is not associated with antioxidant capacity and is independent of pro-vitamin A activity .84 On the other hand, it should be recognized that carotenoids in the epidemiologic studies might have been serving as markers for other protective factors in fruits and vegetables, but were not acting as effective agents themselves. Stimulation of gap junctional communication has been suggested as a possible biochemical mechanism underlying the cancer-preventive activity of carotenoids. It appears that the presence of a six-membered ring substituent at the end of the conjugated system of double bonds is required for the stimulatory effect; five-membered ring carotenoids are less active. However, there is increasing evidence that oxidation products of carotenoids, especially retinoic acid analogs, significantly contribute to this biological property.85 Antioxidants (including carotenoids) have been studied for theirability to prevent chronic disease, since the free radical theory of aging in chronic disease etiology remains pre-eminent. b-carotene and others carotenoids have antioxidant properties, but the antioxidant capability is variable depending on the in vitro system used. The antioxidant activity of these compounds can shift into a prooxidant effect, depending on such factors as oxygen tension or carotenoid concentration. Mixtures of carotenoids alone or in association with others antioxidants can increase their activity against lipid peroxidation.86
A glance of carotenoids economics
Industrially, carotenoids are used in pharmaceuticals, neutraceuticals, and animal feed additives, as well as colorants in cosmetics and foods. The global market for carotenoids can be broken down into 10 products – beta-carotene, lutein, astaxanthin, capsanthin, annatto, canthaxanthin, lycopene, beta-apo-8-carotenal, zeaxanthin, and beta-apo-8-carotenal-ester. According to the study “THE GLOBAL MARKET FOR CAROTENOIDS (FOD025D)” from BCC Research (www.bccresearch.com), the global carotenoids market totaled nearly $1.2 billion in 2010. In 2018, that value is projected to surpass $1.4 billion, increasing at an eight-year compound annual growth rate (CAGR) of 2.3%. The main categories of carotenoids are still the same. There is no, new types of carotenoids have come on the market and, with the exception of astaxanthin, no fundamentally new manufacturing technology has been introduced. However, the market for carotenoids over the past four years has changed dramatically for some product segments. Prices have dropped for a number of products while market value has risen for others. Beta-carotene is still the most prominent carotenoid. While consumption has developed organically, products across all types of formulation and origin have seen a big drop in price due to the massive market entry of Chinese manufacturers with high-quality products. The beta-carotene market is currently oversupplied. Synthetic beta-carotene continues to dominate the market and hopes that fermentation-derived beta-carotene would capture a larger market share has not been fulfilled, with beta-carotene from algae practically disappearing from the market. Lycopene, an antioxidant recommended to prevent prostate cancer, obtained permission from the European Commission (EC) to be used as a food additive and coloring agent years ago, but hopes that such approval would boost the market have not come true. However, consumption in the traditional supplement segment has increased. However, prices have decreased now that many Asian suppliers offer lycopene extracts from tomatoes at acceptable qualities. Consumption of standard astaxanthin, the color of choice for pigmenting fish and shrimp, is expanding in line with the growing aquaculture industry. However, in contrast to other carotenoids, astaxanthin derived from algae is booming as an ingredient for food supplements and prices for such products are skyrocketing. Technological innovations are occurring at an intensity rarely seen during the past decade in the carotenoid sector, ranging from fully controlled tube systems in production to the use of specifically designed microorganisms that can produce astaxanthin in the dark. Canthaxanthin is still the color of choice for providing a red tone in egg yolks and in the pigmentation of salmonid fishes and shrimp. For canthaxanthin, price erosion started early and has not yet been brought to a standstill. Expansion of consumption is moderate and market values are shrinking due to decreasing prices. The lutein supplement market boomed until 2004. Since then, the market has grown at moderate rates. The producer landscape has multiplied, competition among primary lutein producers is fierce, and prices for unrefined feed grade product are still low, although they are mildly increasing. In the supplement segment, lutein enjoys high popularity but prices are declining given the numerous producers in Asia (http://www.reportlinker.com).
Conclusion
Carotenoids are the phytonutrients that impart a distinctive yellow, orange, and red color to various fruits and vegetables. The physico-chemical properties and the biological activities of carotenoids are intimately related to their structures. Amongst several dozen of antioxidants in the foods that we eat, most of these carotenoids have antioxidant activity. Carotenoids have been studied for their ability to prevent chronic disease, since the free radical theory of aging in chronic disease etiology remains pre-eminent. b-carotene, lycopene, zeaxanthine and others carotenoids have antioxidant properties, but the antioxidant capability is variable depending on the in vitro system used. The antioxidant activity of these compounds can shift into a prooxidant effect, depending on such factors as oxygen tension or carotenoid concentration. Mixtures of carotenoids alone or in association with others antioxidants can increase their activity against lipid peroxidation. The isomeric form may be an important factor in the bioavailability and bioactivity of certain carotenoids. Carotenoids as natural pigments, are used by the industry as pharmaceuticals, neutraceuticals, and animal feed additives, as well as colorants in cosmetics and special foods. In many food crops, especially the major staple crops contain only traces to low amounts of carotenoids. Through biotechnology significant progress has been made in developing food crops rich in carotenoids by altering the expression of carotenoids biosynthetic genes or using microorganisms to produce carotenoids by fermentation procedure. The global market for carotenoids can be broken down into 10 products – beta-carotene, lutein, astaxanthin, capsanthin, annatto, canthaxanthin, lycopene, beta-apo-8-carotenal, zeaxanthin, and beta-apo-8-carotenal-ester.
References
- Lampe JW. Health effects of vegetables and fruit: assessing mechanisms of action in human experimental studies. Am J Clin Nutr 1999;70(Suppl):475–90
- Olson, P. James Allen .Recommended dietary intakes (RDI) of vitamin A in humans. Am J Cuin Nut,’ l987;45:704-l6.
- Britton G (1995) Structure and properties of carotenoids in relation to function. Faseb J 9: 1551–1558
- Janik, E.; Grudziński, W.; Gruszecki, W.I.; Krupa, Z. The xanthophyll cycle pigments in Secale cereale leaves under combined Cd and high light stress conditions. J. Photochem. Photobiol. B 2008, 90, 47–52
CrossRef - Niyogi KK: Photoprotection revisited. Annu Rev Plant Physiol Plant Mol Biol 1999, 50:391-417.2.
CrossRef - Niyogi KK: Safety valves for photosynthesis. Curr Opin Plant Biol 2000, 3:455-460.
CrossRef - Andrew D. Parry and Roger Horgan Carotenoids and abscisic acid (ABA) biosynthesis in higher plants. Physiologia Plantarum Volume 82, Issue 2, pages 320–326, June 1991.
- Adrian J. Cutler and Joan E. Krochko. Formation and breakdown of ABA. Trends in plant Sscience , Reviews. December 1999, Vol. 4, No. 12.
- Beecher GR: Nutrient content of tomatoes and tomatoes products.Proc Soc Exp Biol Med 218:98–100, 1998.
CrossRef - Botella-Pavia P and Rodriguez-Concepcion M (2006) Carotenoid biotechnology in plants for nutritionally improved foods.Physiol Plant 126:369–381.
CrossRef - World Health Organization World Health Organization. Global prevalence of vitamin A deficiency in populations at risk 1995−2005; http://whqlibdoc.who.int/publications/2009/9789241598019_eng.pdf (accessed Feb 15,2015)
- Giovannucci E (1999) Tomatoes, tomato-based products, lycopene, and cancer: review of the epidemiologic literature. J Natl Cancer Inst 91:317–331
CrossRef - Hadley CW, Miller EC, Schwartz SJ, Clinton SK (2002).Tomatoes, lycopene, and prostate cancer: progress and promise. Exp. Biol. Med. 227:869–880
CrossRef - Lessin WJ, Catigani GI, Schwartz SJ: Quantification of cis-trans isomers of provitamin A carotenoids in fresh and processed fruits and vegetables. J Agric Food Chem 45:3728–3732, 1997.
CrossRef - Osganian, S. K., Stampfer, M. J., Rimm, E., Spiegelman, D., Manson, J. E., & Willett, W. C. (2003). Dietary carotenoids and risk of coronary artery disease in women. American Journal of Clinical Nutrition, 77(6), 1390-1399. PMid:12791615.
- Gale, C. R., Ashurst, H. E., Powers, H. J., & Martyn, C. N. (2001). Antioxidant vitamin status and carotid atherosclerosis in the elderly. American Journal of Clinical Nutrition, 74(3), 402-408. PMid:11522566.
- 17. Nishino, H., Murakoshi, M., Li, T., Takemura, M., Kuchide, M., Kanazawa, M., Yang Mou, X., Wada, S., Masuda, M., Ohsaka, Y., Yogosawa, S., Satomi, Y., & Jinno, K. (2002). Carotenoids in cancer chemoprevention. Cancer and Metastasis Reviews, 21(3), 257-264. PMid:12549764. http://dx.doi.org/10.1023/A:1021206826750
CrossRef - Nishino, H., Tokuda, H., Murakoshi, M., Satomi, Y., Masuda, M., Onozuka, M., Yamaguchi, S., Takayasu, J., Tsuruta, J., Okuda, M., Khachik, F., Narisawa, T., Takasuka, N., & Yano, M. (2000). Cancer prevention by natural carotenoids. Biofactors, 13(1), 89-94. PMid:11237205. http://dx.doi.org/10.1002/biof.5520130115
CrossRef - Di Mascio, P., Kaiser, S., & Sies, H. (1989). Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Archives of Biochemistry and Biophysics, 274(2), 532-538. http://dx.doi. org/10.1016/0003-9861(89)90467-0
CrossRef - Stahl, W., & Sies, H. (1996). Lycopene: a biologically important carotenoid for humans? Archives of Biochemistry and Biophysics, 336(1), 1-9. PMid:8951028. http://dx.doi.org/10.1006/abbi.1996.
- Stacewicz-Sapuntzakis, M., & Bowen, P. E. (2005). Role of lycopene and tomato products in prostate health. Biochimica et Biophysica Acta, 1740(2), 202-205. PMid:15949687. http://dx.doi.org/10.1016/j. bbadis.2005.02.004.
- Giovannucci, E. (2011). Serum lycopene and prostate cancer progression: a re-consideration of findings from the prostate cancer prevention trial. Cancer Causes Control, 22(7), 1055-1059. PMid:21573862. http://dx.doi.org/10.1007/s10552-011-9776-x
CrossRef - Brasil. Agência Nacional de Vigilância Sanitária. (2008). Lista de alegações de propriedade funcional aprovadas. Brasília: Anvisa. Retrieved from http://www.anvisa.gov.br/alimentos/comissoes/ tecno_lista_alega.htm
- Krinsky NI, Landrum JT, Bone RA (2003) Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu. Rev. Nutr. 23:171–201
CrossRef
- Lorenzo, Y., Azqueta, A., Luna, L., Bonilla, F., Domínguez, G., & Collins, A. R. (2009). The carotenoid β-cryptoxanthin stimulates the repair of DNA oxidation damage in addition to acting as an antioxidant in human cells. Carcinogenesis, 30(2), 308-314. PMid:19056931. http://dx.doi.org/10.1093/carcin/bgn270
CrossRef - Nishi, K., Muranaka, A., Nishimoto, S., Kadota, A., & Sugahara, T. (2012). Immunostimulatory effect of b-cryptoxanthin in vitro and in vivo. Journal of Functional Foods, 4(3), 618-625. http://dx.doi. org/10.1016/j.jff.2012.04.001
CrossRef - Yamaguchi, M. (2012). Role of carotenoid β-cryptoxanthin in bone homeostasis. Journal of Biomedical Science, 19, 36-51. PMid:22471523 PMCid:PMC3361488. http://dx.doi.org/10.1186/1423-0127-19-36
CrossRef - James Allen Olson Absorption, transport, and metabolism of carotenoids in humans Pure &Appl. Chem., Vol. 66, No. 5, pp. 1011-1016, 1994.
- Jalal, F.; Nesheim, M.C.; Agus, Z.; Sanjur, D.; Habicht, J.P. Serum retinol concentrations in children are affected by food sources of β-carotene, fat intake, and anthelmintic drug treatment. Am. J. Clin. Nutr. 1998, 68, 623–629.
- van Het Hof, K.H.; West, C.E.; Weststrate, J.A.; Hautvast, J.G. Dietary factors that affect the bioavailability of carotenoids. J. Nutr. 2000, 130, 503–506.
- Clevidence, B.A.; Bieri, J.G. Asociation of carotenoids with human plasma lipoproteins. In Methods in Enzymology; Abelson, J.N., Simon, M.I., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 33–46.
- Wang, W.; Connor, S.L.; Johnson, E.J.; Klein, M.L.; Hughes, S.; Connor, W.E. Effect of dietary lutein and zeaxanthin on plasma carotenoids and their transport in lipoproteins in age-related macular degeneration. Am. J. Clin. Nutr. 2007, 85, 762–769.
- Kotake-Nara, E.; Nagao, A. Absorption and metabolism of xanthophylls. Mar. Drugs 2011, 9,1024–1037.
CrossRef - Nagao, A. Absorption and metabolism of dietary carotenoids. Biofactors 2011, 37, 83–87.
CrossRef - 35. Khachik, F.; Spangler, C.J.; Smith, J.C., Jr.; Canfield, L.M.; Steck, A.; Pfander, H. Identification,quantification, and relative concentrations of carotenoids and their metabolites in human milk and serum. Anal. Chem. 1997, 69, 1873–1881.
CrossRef - de Pee, S., C.E. West, D. Permaesih, S.M. Muhilal, and J. Hautvast. 1998.Orange fruit is more effective than are dark-green, leafy vegetables in increasing serum concentrations of retinol and b-carotene in schoolchildren in Indonesia. Amer. J. Clin. Nutr. 68:1058–1067.
- Rock CL (1997). Carotenoids: biology and treatment. Pharmacol Ther 75: 185-197
CrossRef - Paetau et al., 1997 Paetau, I., H. Chen, N. Goh, and W.S. White. 1997. Interactions in the postprandial appearance of b-carotene and canthaxanthin in plasma triacylglycerol-rich lipoproteins in humans. Amer. J. Clin. Nutr. 66:1133–1143
- Kostic, D., W.S. White, and J.A. Olson. 1995. Intestinal absorption, serum clearance, and interactions between lutein and b-carotene when administered to human adults in separate or combined oral doses. Amer. J. Clin.Nutr. 62:604–610.
- Micozzi, M.S., E.D. Brown, B.K. Edwards, J.G. Bieri, P.R. Taylor, F. Khachik,G.R. Beecher, and J.C. Smith. 1992. Plasma carotenoid response to chronic intake of selected foods and b-carotene supplements in men. Amer. J. Clin.Nutr. 55:1120–1125.)
- Johnson E.J., J. Qin, N.I. Krinsky, and R.M. Russell. 1997. Ingestion by men of a combined dose of b-carotene and lycopene does not affect the absorption of b-carotene but improves that of lycopene. J. Nutr. 127:1833–1837.
- Boileau, T. W., Boileau, A. C., and Erdman, J. W., Jr. (2002). Bioavailability of all-trans and cis-isomers of lycopene. Exp. Biol. Med. 227: 914–919
CrossRef - Chitchumroonchokchai, C, Schwartz, S. J., Failla, M. L. (2004). Assessment of lutein bioavailability from meals and a supplement using simulated digestion and Caco-2 human intestinal cells. J Nutr. 134: 2280–2286
- Clinton, S. K. (1998).Lycopene:Chemistry, biology, and implications for human health and disease. Nutr Rev. 56(2): 35–51
CrossRef - Kun, Y., Lule, U. S., and Xiao-Lin, A. D. (2006). Lycopene: Its properties and relationship to human health. Food Rev Int. 22: 309–333
CrossRef - Boileau, A. C., Merchen, N. R., Wasson, K., Atkinson, C, and Erdman, J. W.(1999). Cis-lycopene is more bioavailable than trans-lycopene in vitro and in vivo in lymph-cannulated ferrets. J Nutr. 129: 1176–1181
- Boileau, T. W., Boileau, A. C., and Erdman, J. W., Jr. (2002). Bioavailability of all-trans and cis-isomers of lycopene. Exp. Biol. Med. 227: 914–919
CrossRef - Unlu, N. Z., Bohn, T., Francis, D., Clinton, S. K., and Schwartz, S. J. (2007).Carotenoid absorption in humans consuming tomato sauces obtained from tangerine or high-beta-carotene varieties of tomatoes. J Agr Food Chem. 55:1597–1603.
CrossRef - Boileau, A. C., Merchen, N. R., Wasson, K., Atkinson, C, and Erdman, J. W.(1999 Cis-lycopene is more bioavailable than trans-lycopene in vitro and in vivo in lymph-cannulated ferrets. J Nutr. 129: 1176–1181.
CrossRef - Levin, G., Mokady, S. (1995). Incoporation of all-trans- or 9-cis-beta-carotene into mixed micelles in vitro. Lipids. 30(2):177–179
- Ferruzzi, M. G., Lumpkin, J. L., Schwartz, S. J., and Failla, M. L. (2006).Digestive stability, micellarization, and uptake of beta-carotene isomers by Caco-2 human intestinal cells. J Agr Food Chem. 54(7): 2780–2785.
CrossRef - Dewanto, V., Wu, X., Adorn, K. K., and Liu, R. H. (2002). Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J Agr Food Chem. 50(10): 3010–3014
CrossRef - Schierle, J., Bretzel, W., Buhler, I., Faccin, N., Hess, D., and Steiner, K., and Schuep, W. (1997). Content and isomeric ratio of lycopene in food and human blood plasma. Food Chem. 59(3): 459–465
CrossRef - Stahl,W., and Sies, H. (1992). Uptake of lycopene and its geometrical isomers is greater from heat-processed than from unprocessed tomato juice in humans.J Nutr. 122: 2161–2166.
- Stahl, W., and Sies, H. (1996). Lycopene: A biologically important carotenoid for humans? Arch Biochem Biophys. 336(1):1–9
CrossRef - Agarwal, A., Shen, H., Agarwal, S., and Rao, A. V. (2001). Lycopene content of tomato products: Its stability, bioavailability and in vivo antioxidant properties. J. Med. Food. 4(1): 9–15.
CrossRef - Xianquan, S., Shi, J., Kakuda, Y., and Yueming, J. (2005). Stability of lycopene during food processing and storage. J Med Food. 8(4):413–422.)
CrossRef - Schieber, A.; Carle, R. Occurrence of carotenoid cis-isomers in food: Technological, analytical,and nutritional implications. Trends Food Sci. Technol. 2005, 16, 416–422.
CrossRef - Marx Michaela, Stuparic Monika, Schieber Andreas and Reinhold Carle Effects of thermal processing on trans–cis-isomerization of b-carotene in carrot juices and carotene-containing preparations. Food Chemistry 83 (2003) 609–617
CrossRef - 60. Li, Li and Ζ Joyce Van Eck. Metabolic engineering of carotenoids accumulation by creating a metabolic sink. Transgenic Res. (2007) 16:581–585)
- Ribeiro, B.D.; BARRETO, D.W. and COELHO, M.A.Z. (2011). Technological aspects of β-carotene production. Food and Bioprocess Technology, vol. 4, no. 5, p. 693-701
CrossRef - Paz, E.D.; MARTÍN, Á. ESTRELLA, A.; RODRÍGUEZ-ROJO, S.; MATIAS, A.A.; DUARTE, C.M.M. and Cocero, M.J. (2012). Formulation of β-carotene by precipitation from pressurized ethyl acetate-on-water emulsions for application as natural colorant. Food Hydrocolloids, vol. 26, no. 1, p. 17-27
CrossRef - Gautam, S.; PLATEL, K. and SRINIVASAN, K. (2010). Influence of β-carotene-rich vegetables on the bioaccessibility of zinc and iron from food grains. Food Chemistry, vol. 122, no. 3, p. 668-672
CrossRef - Mapari, S.A.S.; THRANE, U. and MEYER, A.S. (2010). Fungal polyketide azaphilone pigments as future natural food colorants? Trends in Biotechnology, vol. 28, no. 6, p. 300-307.
CrossRef - Mogedas, B.; CASAL, C.; FORJÁN, E. and VÍLCHEZ, C. (2009). β-Carotene production enhancement by UV-A radiation in Dunaliella bardawil cultivated in laboratory reactors. Journal of Bioscience and Bioengineering, vol. 108, no. 1, p. 47-51.
CrossRef - Mortensen, A. (2006). Carotenoids and other pigments as natural colorants. Pure and Applied Chemistry, vol. 78, no. 8, p. 1477-1491
CrossRef - Malisorn and Suntornsuk, 2009; MOGEDAS, B.; CASAL, C.; FORJÁN, E. and VÍLCHEZ, C. (2009). β-Carotene production enhancement by UV-A radiation in Dunaliella bardawil cultivated in laboratory reactors. Journal of Bioscience and Bioengineering, vol. 108, no. 1, p. 47-51.;
CrossRef - Filotheou, A.; NANOU, K.; PAPAIOANNOU, E.; ROUKAS, T.; KOTZEKIDOU, P. and Liakopoulou Kyriakides, M. (2012). Application of response surface methodology to improve carotene production from synthetic medium by Blakeslea trispora in submerged fermentation. Food and Bioprocess Technology, vol. 5, no. 4, p. 1189-1196
CrossRef - Haila, K. M., Lievonen, S. M., and Heinonen, M. I. (1996). Effects of lutein, lycopene, annatto, and gamma-tocopherol on autoxidation of triglycerides.Journal of Agricultural and Food Chemistry 44(8):2096–2100.
CrossRef - Heinonen, M., Haila, K., Lampi, A. M., and Piironen, V. (1997). Inhibition of oxidation in 10% oil-in-water emulsions by beta-carotene with alpha and gamma-tocopherols. Journal of the American Oil Chemists Society 74(9):1047–1052.
CrossRef - Ribeiro, H. S., Ax, K., and Schubert, H. (2003). Stability of lycopene emulsions in food systems. Journal of Food Science 68(9):2730–2734
CrossRef - Rodriguez-Huezo, M. E., Pedroza-Islas, R., Prado-Barragan, L. A., Beristain,C. I., and Vernon-Carter, E. J. (2004). Microencapsulation by spray drying of multiple emulsions containing carotenoids. Journal of Food Science 69(7):E351–E359
CrossRef - Naves, M.M.V., Moreno, F.S., 1998. Beta-carotene and cancer chemoprevention: from epidemiological association to cellular mechanisms of action. Nutrition Research 18 (10), 1807–1824.
CrossRef - Omenn, G.S., Goodman, G.E., Thornquist, M.D., 1996. Effects of a combination of beta-carotene and vitamin A on lung cancer and cardiovascular disease. New England of Journal of Medicine 334, 1150–1155
CrossRef - Ribeiro, H.S., Cruz, R.C.D., 2005. Biliquid foams containing carotenoids. Engineering in Life Sciences 5, 84–88
CrossRef - Orset, S., Leach, G.C., Morais, R., Young, A.J., 1999. Spray-drying of the microalga Dunaliella salina: effects on beta-carotene content and isomer composition. Journal of Agricultural and Food Chemistry 47, 4782–4790. ;
CrossRef - Rodriguez-Huezo, M.E., Pedroza-Islas, R., Prado-Barragan, L.A., Beristain, C.I., Vernon-Carter, E.J., 2004. Microencapsulation by spray drying of multiple emulsions containing carotenoids. Journal of Food Science 69, 351–359
CrossRef - Jochen, W., Paul, T., McClements, D.J., 2006. Functional materials in food nanotechnology. Journal of Food Science 71 (9), 107–116.
CrossRef - Quintanilla-Carvajal, M.X., Camacho-Díaz, B.H., Meraz-Torres, L.S., Chanona-Pérez, J.J., Alamilla-Beltrán, L., Jimenéz-Aparicio, A., Gutiérrez-López, G.F., 2010. Nanoencapsulation: a new trend in food engineering processing. Food Engineering Reviews 2, 39–50.
CrossRef - Halliwell B. Oxidative stress, nutrition, and health. Experimental strategies for optimization of nutritional antioxidant intake in humans, Free Radical Res. 25 (1996) 57–74. ,
CrossRef - Davies, K.J.. Oxidative stress: the paradox of aerobic life, Biochem. Soc. Symp. 61 (1995) 1–31.
CrossRef - Mathews-Roth, M.. Recent progress in the medical applications of carotenoids, Pure Appl. Chem. 63 (1991) 147–156.
CrossRef - Rosangela Novo, Paula S. Azevedo, Marcos F. Minicucci, Leonardo A. M. Zornoff, Sergio A. R. Paiva. Effect of Beta-Carotene on Oxidative Stress and Expression of Cardiac Connexin 43. Arq Bras Cardiol. 2013 September; 101(3): 233–239. doi: 10.5935/abc.20130160.
CrossRef - Bendich A: Biological functions of dietary carotenoids. Ann.N.Y. Acad. Sci. 691:61–67, 1993.
CrossRef - Sies H, Stahl W: Carotenoids and intercellular communication via gap junctions. Int J Vitam Nutr Res 67:364–367, 1997
- McNulty ,Hyesun P. , Jungsoo Byun , Samuel F. Lockwood , Robert F. Jacob , R. Preston Mason a Differential effects of carotenoids on lipid peroxidation due to membrane interactions: X-ray diffraction analysis. Biochimica et Biophysica Acta 1768 (2007) 167–174.
CrossRef - Southon S, Faulks R (2003). Carotenoids in food: bioavailability and functional benefits. In: Phytochemical functional foods. Johnson, I and Williamson, G (Eds.). Ch. 7. Woodhead Publishing Limited. CRC Press. ISBN 0-8493-1754-1, pp. 107-127.
CrossRef





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








