
Introduction
Anthocyanins found in plant material or their fruits have been widely consumed in various geographical regions. Anthocyanins are water-soluble plant-derived subgroups of flavonoids accountable for the pigmentation of red, purple, and blue, also present in a glycosylated form exhibiting antioxidant activities, a potential dietary component for nutritional management in various health disorder1,-4. The term anthocyanin is a fusion of two Greek words: anthos, which means “flowers,” and kyanos, that means “dark blue”5. Common anthocyanins include pelargonidin, cyanidin, peonidin, delphinidin, petunidin, and malvidin6.
Anthocyanins are prevalent in blue and red vegetables and fruits, with their concentrations in plants significantly differing among species, influenced by variety or cultivar, geographic location, climate, agricultural practices, harvest timing, processing, seasonal fluctuations, storage conditions, ripening, temperature, and light exposure7. Blue, red, and purple berries, plums, apples, grapes, cherries, cabbage, and foods with high concentrations of organic colourants are dietary sources of anthocyanins8. Glycosylation makes anthocyanins more soluble and bioavailable. Additionally, it affects their antioxidant activity and possible health advantages by influencing how they are Incorporated, mobilized and utilized in the body. Acetylation improves anthocyanin stability, particularly in the face of environmental stressors such as light, heat, and pH fluctuations. Their biological activity may be prolonged as a result of their increased pigment efficacy and resistance to deterioration. These alterations improve the biological and functional qualities of anthocyanins, including their antioxidant and anticancer capabilities8.
According to a number of studies, anthocyanins have strong antioxidant qualities that allow them to counteract the body’s dangerous free radicals and shield cells from oxidative damage9,-12. Anthocyanins contain anti-inflammatory properties that might be useful in the prevention and management of chronic inflammatory illnesses13-15. Epidemiological studies indicate that anthocyanin intake reduces the risk of diabetes, arthritis, cardiovascular disease, and cancer, attributable to their antioxidant and anti-inflammatory effects16-19. Anthocyanins also benefit eye health, particularly when it comes to issues caused by diabetic retinopathy — high serum blood glucose levels and eye-related disorders macular degeneration20, 21. Research shows that ingesting foods high in anthocyanins may boost cognitive function, improve memory, and have a beneficial influence on several neurodegenerative illnesses, which is especially significant in the context of an aging population22, 23.
There are several research papers explaining the role of anthocyanins as anti-inflammatory and anti-cancer as a whole. Comprehensive study on dose dependent cell line and clinical trial related to molecular and functional aspect as antioxidant and anti-inflammatory still not clear. Functional pathway including dosage and duration may help to optimize maximum level of anthocyanin consumption for achieving optimal health benefits. This study was particularly conducted to explore their molecular and functional aspects, with a focus on their mechanisms related to antioxidant and anti-inflammatory actions. This article will examine the primary origin of anthocyanins, the mechanisms by which they particularly act on inflammation and cancer evaluating the most recent scientific research including in-vitro and in-vivo evidence supporting the role of anthocyanins in modulating oxidative stress, inflammation, and cancer prevention. Our primary focus will be anthocyanins and their subcomponents consumption in association with inflammation and anticancer activities in scientific studies.
Materials and Methods
We conducted a literature search on the effect of anthocyanins on inflammation and cancer. The research was carried out by searching for specific keywords, such as “Anthocyanins, delphinidin, cyanidin, anticancer, anti-inflammatory, immunomodulation” on search engines Pubmed and Google Scholar. Literature published from the years 2013 to 2024 was included with primary focus on studies employing human cell culture model and human clinical trials. The first selection was based on title and abstract screening. Studies reporting specific outcomes such as a dose-dependent decrease in ROS production, induction of early apoptosis in cancer cell lines, promotion of endothelial cell permeability, modulation of cell-cell adhesion and facilitation of trans endothelial migration and activation of the antioxidant defence system were included. Studies with an experimental placebo control are also considered for inclusion. Animal studies involving rodents and Drosophila are excluded from this review. Studies using anthocyanins in combination with other polyphenols are also excluded. Research lacking a control group is not considered. Studies published in languages other than English as well as dissertations, review articles and book chapters are excluded from this review. Some publications were discarded owing to inconsistencies with the subject, or generic material that did not specifically address the connection under study. After analyzing and reviewing the extracted research articles, the data was compared for reliability.
Results
Chemical structure
Anthocyanins are flavonoids that give vegetables, flowers, and fruits their blue, red, and purple, colours24. They provide several health advantages and protect plants from environmental hazards. Plant pigments are present in vacuoles, often in fruits and flowers as granules ranging from 3 to 10 µm in diameter25. Anthocyanins are glycosides composed of a sugar molecule an aglycone (anthocyanidin)26. Their chemical structure determines their biological qualities and colour. Anthocyanins are composed of three rings: two aromatic rings (A and B) and one heterocyclic ring with oxygen (C) of cationic nature. They frequently exist as flavylium (2-phenylchromene). The C15 skeleton is composed of a chromate ring that contains a second aromatic ring B in position 2 (C6-C3-C6). The flavylium cation is the basic structure of anthocyanidins27. The model structure is shown in Figure 1.
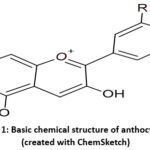 |
Figure 1: Basic chemical structure of anthocyanins (created with ChemSketch) |
Anti-inflammatory and antioxidant effect Anthocyanin
The free radical scavenging capacity of anthocyanins is significantly affected by their chemical structure, and not all of them exhibit comparable activities in scavenging a variety of Reactive Nitrogen Species (RNS) and ROS. The antioxidant capacity of anthocyanins is contingent upon the basic structural orientation of the compound, as the ring orientation determines the ease with which a hydrogen atom from a hydroxyl group can be donated to a free radical and the anthocyanin’s ability to support an unpaired electron28. The degree and position of hydroxyl groups in the B ring, together with the degree and location of methoxyl groups, affect the stability, reactivity, and subsequently the antioxidant properties of anthocyanins29. NF-κB cells are essential in the regulation of genes associated with immunity and inflammation. It has been shown to enhance the invasive phenotype and transcription of oncogenes in cancer cells30. Free radicle scavenging and inflammation modulation are the mechanisms of anthocyanins for cancer prevention.
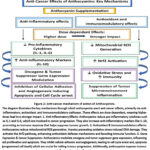 |
Figure 2: Anti-cancer mechanism of action of Anthocyanins. |
The diagram illustrates the key mechanisms through which anthocyanins exert anti-cancer effects, primarily via anti-inflammatory, antioxidant, and immunomodulatory pathways. These effects are dose-dependent, meaning higher doses lead to a stronger impact.1. Anti-Inflammatory Effects: Anthocyanins reduce pro-inflammatory cytokines such as IL-1 and IL-6, which are involved in cancer progression. They also increase anti-inflammatory markers like IL-10, promoting an immune response that helps suppress tumour growth. 2. Antioxidant & Immunomodulatory effects: Anthocyanins reduce mitochondrial ROS generation, thereby preventing oxidative stress-induced DNA damage. They activate the Nrf2 pathway, enhancing antioxidant defense mechanisms and boosting immune function. 3. Gene Expression & Cancer Suppression: Anthocyanins modulate oncogene and tumor suppressor gene expression, influencing cell proliferation and apoptosis. They inhibit cellular adhesion and angiogenesis, leading to cell cycle arrest and, apoptosis (programmed cell death) which are crucial for halting tumour progression. Additionally, anthocyanins suppress tumour microenvironment inflammation, a key factor in cancer metastasis.
Increasing evidence from randomized controlled trials (RCTs) and mechanistic studies confirms anthocyanins’ potential to prevent oxidative stress and inflammation by modulating important signaling pathways, such as Nrf2 and NF-κB. Current study evaluates the effects of anthocyanins on oxidative stress, inflammation, and metabolic health using both in-vitro and in-vivo models. The findings are categorized into cellular responses, and human trials, which explore anthocyanin supplementation’s impact on human health. The key outcomes focus on anthocyanins’ ability to modulate inflammatory signaling, enhance antioxidant defences, regulate lipid metabolism, and improve cardiovascular function.
Table 1: Summary of antioxidant and anti-inflammatory effect of Anthocyanin Extracts on cell cultures.
|
Model |
Intervention | Findings | Reference |
| TNFα induced Endothelial cell | A mixture of Cy3Gal, C3G, cyanidin-3-arabinoside, D3G, P3G and 4-hydroxybenzaldehyde | ↓trans endothelial migration, monocyte adhesion and
modulation of genetic expression |
31 |
|
HaCat keratinocytes cell line
|
Hibiscus syriacus L extract
|
↓H2O2-mediated apoptosis, ↓mitochondrial ROS generation, maintaining mitochondrial membrane potential, and activating Nrf2 | 32 |
| Caco-2 intestinal cells
|
Wild blueberry extract
|
IL-1β-induced NF-κB activation in intestinal epithelial and ↓Caco-2 cells dose-dependently
|
33 |
|
Cancer cell lines |
Black rice extract (P3G + C3G)
|
Dose-dependent ↓cellular proliferation.
↓Apoptosis in cell lines. |
34 |
| Oxidant model | Purple rice extract
|
↓Inflammation in chondrocytes via ↓NF-κB signalling |
35 |
|
Cancer cell line |
Purple tea shoots extract | Expression of apoptotic fragment formation and nuclear compaction in affected cells.
Induction in mononuclear cell proliferation |
36 |
| Cancer cell line | L. caerulea ‘Beilei’ fruit extract | Induce late apoptosis to early apoptosis.
↓Cell division G2 /M phase differentiation and proliferation |
37 |
|
Oxidant model |
C3G and D3G | Protect GI physiology in HFD-induced obesity
↓Caco-2 monolayer permeabilization, ↑NOX1/4, and NF-κB/ERK activation caused by tumor necrosis factor alpha. |
38 |
|
Cancer cell line |
C3G and D3G | ↓Expression of the PD-L1 protein in HCT-116 cells and the intensity of PD-L1 fluorescence by 39%.
Induce apoptosis in HTC cell lines. |
39 |
| Cancer cell line | Extract from strawberries | ↓Triglycerides and LDL-cholesterol and ↑HepG2 cell redox status |
40 |
|
High glucose- induced human retinal capillary endothelial cells |
Extract from blueberry | ↓ROS production, by ↑SOD activity.
↓VEGF expression and Akt pathway
|
41 |
| A549-lung cells and THP-1 macrophages | Extract from black pigmented rice | ↓SP-induced inflammation by ↑NF-kB and ↓inflammasome-dependent inflammatory pathway proteins (NLRP3, ASC, and capase-1). |
42 |
|
Epithelial cell |
C3G | ↓TNF-α-induced nuclear translocation of NF-κB and IL-8 gene expression in Caco-2 cells. | 43 |
| Cancer cell line | Fermented grape and strawberry extract | ↑Apoptosis by regulation of the Caspase-3 pathway in HT-Colon cancer cells. |
44 |
|
Endothelial cells |
Blueberry lyophilized extract | ↑EC proliferation, a key step in angiogenesis and wound healing. |
45 |
|
Infection model hepatitis C virus (HCV) |
Delphinidin | ↓Viral envelope inhibition of Hepatitis C entry. |
46 |
|
Oxidative cell model |
Native Chilean Berries Extract | A dose-dependent ↓oxidation | 47 |
| Whole blood culture with LPS | Pelargonidin-3-O-glucoside and its metabolites. | No significant activity oxidative burst |
48 |
Endothelial and Keratinocyte Models
Anthocyanins have demonstrated significant anti-inflammatory and antioxidant effects in various cellular models (Table 1). In a TNF-α-stimulated endothelial cell model, a mixture of anthocyanin extracts (Cy3Gal, C3G, cyanidin-3-arabinoside, D3G, P3G, and 4-hydroxybenzaldehyde) inhibited trans-endothelial migration and monocyte adhesion. The study found a modulation of gene expression in cell-cell adhesion mechanisms, confirming anthocyanins’ protective role in vascular health31. In an oxidative stress model using HaCat keratinocyte cell lines, anthocyanins extracted from Hibiscus syriacus L. reduced H₂O₂-mediated apoptosis by decreasing mitochondrial ROS production, stabilizing mitochondrial membrane potential, and activating the Nrf2 pathway, which upregulated antioxidant defence genes such as HO-1 and NQO1. This mechanism provided strong protective effects against oxidative damage in skin cells32. A32n IL-1β-induced oxidative stress model in Caco-2 cells demonstrated that anthocyanin-rich wild blueberry extract suppressed NF-κB activation thereby mitigating gut inflammation and oxidative stress in intestinal epithelial cells dose depentdently33. Blueberry anthocyanins inhibited VEGF expression, Akt pathway and upregulated SOD activity, contributing to oxidative stress reduction on high glucose- induced human retinal capillary endothelial cells 41.
Cancer Cell Line Studies
Anthocyanins have also been studied in cancer models. P3G and C3G induced a dose-dependent inhibition of cellular proliferation and apoptosis in multiple cancer cell lines, demonstrating their potential as therapeutic agents34. Anthocyanins extracted from purple rice reduced inflammation by suppressing NF-κB signaling in chondrocytes, indicating potential benefits for joint health and arthritis management35. Anthocyanins from purple tea shoots induced apoptotic fragment formation and nuclear compaction, while L. caerulea ‘Beilei’ fruit extract (100 μg/mL) promoted G2/M phase arrest, significantly inhibiting cancer cell proliferation36,37. D3G and C3G induced apoptosis in HTC cell lines. It also reduced expression of the PD-L1 protein in HCT-116 cells and the intensity of PD-L1 fluorescence by 39%39. Anthocyanins from strawberries reduced triglycerides and LDL-cholesterol and enhanced HepG2 cell redox status via modifying thiobarbituric acid-reactive compounds, antioxidant enzyme activity, and ROS formation40.
Gut, Hepatic, and Immune Models
Additional studies have explored the broad antioxidant and anti-inflammatory effects of anthocyanins. Anthocyanins prevented Caco-2 monolayer permeabilization, NOX1/4 upregulation, oxidative stress, and NF-κB/ERK activation caused by tumor necrosis factor alpha and Protect GI physiology in HFD-induced obesity via redox-regulated pathways.38 C3G suppressed NF-κB activation and reduced p65 nuclear translocation in macrophage cell lines, leading to reduced inflammatory responses42. C3G also downregulated pro-inflammatory proteins while upregulating anti-inflammatory markers, further validating its anti-inflammatory role43. Fermented grape and strawberry anthocyanins were shown to induce caspase-3-mediated apoptosis in colon cancer cells, supporting their potential anticancer properties44. A blueberry lyophilized extract significantly reduced inflammatory cell counts and regulated phospholipase C pathway gene expression, providing further evidence of anthocyanins’ anti-inflammatory effects45. In an oxidative stress-induced hepatocyte model, anthocyanins from native Chilean berries exhibited a dose-dependent decrease in oxidative stress markers, confirming hepatoprotective effects47. Delphinidin significantly inhibited HCV viral envelope formation, suggesting potential antiviral applications of anthocyanins46. In another study in cellular models, Pelargonidin-3-O-glucoside and its metabolites showed no significant activity on oxidative stress48.
Table 2: Summary of antioxidant and anti-inflammatory effect of Anthocyanin on human trials.
|
Population |
Intervention with duration | Findings | Reference |
| Healthy Young Adults | Extracted Anthocyanins
Duration of 14 days |
Dose-dependent ↓8-iso prostaglandin and plasma interleukin-10 F2α. |
49 |
|
Healthy Adults |
Purified Anthocyanins
Duration: 5h |
Modulate the associated postprandial dysmetabolism, endotoxemia, alterations of glycemia and lipidaemia, and redox and insulin signalling. | 50 |
| Adults with Dyslipidaemia | Purified Anthocyanins
Duration: 6 weeks and 12 weeks |
Dose-response association with
↓8-OHdG, IL-6, 8-iso-PGF2α, MDA, TNF-α, and oxidative stress biomarkers |
51 |
|
Older adults BMI above 30.6 kg/m2
|
High-fat high-energy meal with anthocyanins-rich queen garnet plum
Duration: 2 h |
↓CRP levels, ↓IL-6. | 52 |
| Post menopausal women | Elderberry extracts
Duration: 12 weeks |
No significant impact on fatty acid profiles and biomarkers of CVD. |
53 |
|
Normal healthy and MetS (age 25-75y) male and female |
Purified anthocyanins
Duration: 4 weeks |
Significant ↓Uric acid, hs-CRP, FBG, HDL, LDL, TG, Cholesterol | 54 |
| Endurance sport athlete | New Zealand blackcurrant powder
Duration for 7 days |
↓Total peripheral resistance by 16%, improved stroke volume and cardiac output by 25% and ↓plasma lactate |
55 |
|
Healthy Adults |
Tart cherry powder.
Duration: 2-week placebo control trials |
No notable drop-in heart rate, and diastolic blood pressure, deoxygenated hemoglobin | 56 |
| Pre hypertensive adults | Anthocyanins
Duration: 4 weeks
|
↑HDL cholesterol production in pre-hypertensive and non-dyslipidaemia patients. |
57 |
|
Adults with mild cognitive impairment |
Anthocyanins extract
Duration: 8 weeks |
↓Serum TNF-α in senior adults. No effects noted on other inflammatory biomarkers (IL-6, hs-CRP, and IL-1β), microcirculation function, or 24-hour ambulatory blood pressure. No effects noted in the group that received low dose of food anthocyanins. |
58 |
|
Healthy age male |
New Zealand blackcurrant extracts
Duration: 14 days |
Longer duration of intake ↑fatty acid oxidation and alters substrate oxidation metabolism in moderate-intensity exercises. | 59 |
| Brazilian Adults
BMI between 30 and 39
|
Jucara freeze dried pulp
Duration: 6 weeks
|
↓MCP-1 and IL-6 TNF-α production, ↑IL-10 mRNA concentration, and ↓pro-inflammatory condition in obesity. |
60 |
|
Adult with T2DM |
Anthocyanins
Duration: 4 weeks |
Significant ↓IL-18, IL-6, and TNF- α observed in T2DM group. | 61 |
| Men and postmenopausal women aged 18 to 50 years, Sedentary population. | Blueberry powder
Duration: 6 weeks |
↓NK cells, AIx, ASP, and diastolic pressures in sedentary males and females. |
62 |
|
Age group 25 – 60 yrs with a history of hypertension and obesity, on nitrate or high dosage statin medications (rosuvastatin > 20 mg/day, atorvastatin > 40 mg/day or) |
Dried purple-black barberry along with sucrose
Duration for 8 weeks |
↓Lipid profile (TC: LDL-C, HDL-C, sd-LDL-C and TG): Plasma CRP levels in the barberry group in comparison to the placebo group, and a tendency toward ↓plasma IL-6 levels was noted in the barberry group. |
63 |
Systemic Anti-inflammatory and Oxidative Stress Modulation
A 14-day randomized controlled trial in healthy adults demonstrated a dose-dependent decrease in 8-iso prostaglandin F2α and IL-10 levels with anthocyanin intake of 20–320 mg/day, with significant effects observed at doses above 80 mg/day49. A 6–12week randomized crossover trial in dyslipidaemia patients receiving 40–320 mg purified anthocyanins daily showed significant reductions in oxidative stress markers, including 8-OHdG, IL-6, 8-iso-PGF2α, MDA, and TNF-α51.
Cardiovascular and Metabolic Benefits
A single 5-hour dose of 320.4 mg anthocyanins significantly improved postprandial metabolism, reducing endotoxemia, glycemia, and lipidemia50. A 12-week study in postmenopausal women consuming 500 mg of elderberry extract daily found no significant effects on fatty acid profiles53. In a 4-week trial, 80 mg anthocyanins significantly lowered uric acid, hs-CRP, fasting blood glucose, HDL, LDL, triglycerides, and cholesterol levels54. Athletes taking 6 g/day of New Zealand blackcurrant powder for 7 days showed a 25% increase in cardiac output and stroke volume, with reduced plasma lactate accumulation55. A study on tart cherry powder (400–480 mg for 2 weeks) showed a reduction in deoxygenated hemoglobin, although heart rate remained unchanged56. A study in hypertensive patients with metabolic disorders supplemented with 250 mg anthocyanins daily for 8 weeks resulted in reduced IL-6 and TNF-α levels58. A study in pre-hypertensive adults who consumed 640 mg of anthocyanins daily for 4 weeks observed a significant increase in HDL cholesterol57. Another study in sedentary postmenopausal women who consumed 250 g of blueberries daily for 6 weeks reported improved NK cell activity, decreased arterial stiffness, and lowered diastolic blood pressure62. A study in hypertensive patients (BMI >30 kg/m²) consuming 10 g of dried purple-black barberry for 8 weeks demonstrated improvements in lipid profiles and reduced CRP levels63.
Dose-Response Relationship and Optimal Duration
A meta-analysis determined that anthocyanin doses greater than 200 mg/day for 4 weeks significantly enhanced vascular function and reduced inflammatory markers, but intake beyond 500 mg/day did not proportionally increase benefits, suggesting a saturation threshold52. Another study found that short-term high-dose treatments provided better outcomes than long-term low-dose supplementation59. A study in obese individuals found that 5 g/day of freeze-dried Jucara pulp for 6 weeks led to increased IL-10 expression and reduced pro-inflammatory markers60. A study in Type 2 Diabetes Mellitus patients receiving 320 mg/day of anthocyanins for 4 weeks showed significant reductions in IL-6, IL-18, and TNF-α61.
Discussion
A harmful factor’s effect causes the body to respond widely, which is known as inflammation. The factor may be mechanical, chemical, biological, or physical. Additionally, the causes may be separated into external and endogenous categories. Inflammation serves to mobilize the body, cells, and regulators that the cells create to combat and neutralize the danger. Redness, inflammation, fever, impairment of function, and discomfort are the hallmarks of inflammation64,65. Cancer also can be caused by a variety of factors, including genetics (e.g., BRCA2 and BRCA1 in breast cancer), environmental exposure to oncogenic agents (e.g., tobacco smoke, asbestos, UV radiation), environmental influences (e.g., low-vegetable diet, lack of physical activity, excessive alcohol consumption, smoking), and infections (e.g., human papillomavirus HPV, hepatitis B)66,67. According to recent research, anthocyanins, natural plant pigments are gaining popularity because of their many health advantages, which include their ability to reduce inflammation as well as cancer. Anthocyanins may be useful in the management and avoidance of inflammation because of their potent antioxidant qualities. Thus, they show also anticancer effects.
The research revealed that the sources of anthocyanin varied. Diverse fruits and rice were used as an intervention; the fruits used were purple black barberries63, blueberries33,41,45,62 jucara pulp60, black currant extracts55,59, chilean berries47, fermented strawberry and grape44, tart cherry56, elderberry53, and queen garnet plum52 and the rice are black rice and purple rice34-35,42. Certain studies also made use of purified anthocyanins. Among the studies, monomeric anthocyanin components were used, including pelargonidin-3-O-glucoside, D3G, and C3G44,46,48. Orally administered anthocyanins in the form of supplementation and/or by addition to diet or related to the intake of food included in the mentioned studies anthocyanin intervention ranges widely from 1 week to 8 weeks49-51.
Among all the study publications, some investigated a decrease in ROS activity35,45; moreover, studies on cancerous cells revealed that anthocyanin intervention had beneficial effects. Some studies showed the stimulation of cellular redox pathways and investigations in which anthocyanin intervention had no meaningful effects48. Research including all research indicated that Nrf2 decreased the formation of ROS40,41. This includes research that observed an elevation in catalase activity40, an enhancement in SOD activity41, encourage EC proliferation, a key step in angiogenesis and wound healing45 and a study that noted an increase in caspase activity44. One study indicated an increase in HDL cholesterol among pre-hypertensive and non-lipidemic individuals, an enhancement in fatty acid oxidation during moderate-intensity exercise, and an increase in interleukin-10 mRNA concentration57.
Dose dependent result observed in clinical trials. Dietary intake of anthocyanin extracts showed dose-dependent decrease in 8-iso prostaglandin F2α and IL-10 levels and significant reductions in oxidative stress markers49,51,58. Dosage 300-320 mg anthocyanin extracts showed optimal results47,49, 51, 54, 61; A dose-dependent relation was witnessed higher doses are more effective in short duration54,55.
The findings of the studies under consideration demonstrate how anthocyanin extracts have anti-inflammatory and antioxidant properties when applied to cell cultures. In one study, anthocyanin extracts were shown to modify gene expression associated with cell-cell adhesion by drastically reducing monocyte adherence and trans endothelial migration in endothelial cells exposed to TNF-α31. Another study on HaCat keratinocyte cell lines demonstrated that anthocyanin extracts from Hibiscus syriacus L. alleviated oxidative stress-induced apoptosis by stabilising mitochondrial membrane potential and decreasing ROS production32. Furthermore, anthocyanins activated the Nrf2 pathway, initiating antioxidant defence mechanisms, thereby reinforcing their protective function against inflammation and oxidative stress32. The findings indicate that anthocyanins, regardless of dosage and duration, produce beneficial effects via multiple cellular pathways, highlighting their potential for therapeutic applications. Additional research is required to establish the maximum level of anthocyanin consumption for achieving optimal health benefits while maintaining safety. Metabolic variability among individuals, as a function of genetic polymorphisms and gut microbiota, can potentially be important in establishing dosage recommendations for optimal effects. The interaction of anthocyanins with food components and their sustained effects over the long term need to be factored into future intervention studies.
Conclusions
This narrative review evaluates the effects of anthocyanins in the form of extracts or association with other foods and their impact as antioxidant and anti-inflammatory component. Therefore, this narrative review verifies that anthocyanins from different food sources improve inflammatory parameters. This study addresses the crucial function of anthocyanins in regulating inflammatory and oxidative stress pathways, demonstrating their potential therapeutic benefits in chronic diseases. The findings demonstrate that anthocyanin supplementation substantially reduces pro-inflammatory cytokines, like IL-1 and IL-6 while increasing anti-inflammatory markers like Interleukin-10. The review also emphasizes how the bioactivity of anthocyanins is dose-dependent, with greater impacts being generated by larger quantities given for a shorter period. Mechanistic discoveries show that anthocyanins have anti-cancer capabilities because they affect the manifestation of genes associated with endothelial function and cellular adhesion. In addition, anthocyanins are further demonstrated as potent antioxidants with immunomodulatory potential by regulating the generation of mitochondrial ROS and stimulating Nrf2. Therefore, the research offers strong evidence in favour of including dietary interventions high in anthocyanins to reduce oxidative stress and inflammation-related illnesses.
However, present data firmly supports anthocyanins’ anti-inflammatory and anti-cancer characteristics. Further study is required to completely understand the mechanisms of action and optimize anthocyanins usage for the prevention of inflammation including cancer.
Acknowledgment
Authors are thankful to Symbiosis Skills and Professional University for providing literature collection facilities. We acknowledge that all listed authors have made a significant scientific contribution to the research in the manuscript approved its claims and agreed to be an author.
Funding Sources
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
Even though adequate data has been given in the form of tables and figures, however, all authors declare that if more data required then the data will be provided on request basis.
Ethics Approval Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to Reproduce Material from Other Sources
Not applicable
Clinical Trial Registration
This research does not involve any clinical trials.
Author Contributions
- Anindita Ghosh: Conceptualization, Methodology, Writing-original draft
- Sunanda Biswas: Visualization, Supervision, Writing – Review & Editing
- Sneha De: Methodology, Writing-original draft
- Samyak Ingale: Methodology, Data collection, Analysis, Writing-original draft
References
- Emamat H., Zahedmehr A., Asadian S., Nasrollahzadeh J. The effect of barberry (Berberis integerrima) on lipid profile and systemic inflammation in subjects with cardiovascular risk factors: A randomized controlled trial. BMC Complementary Medicine and Therapies. 2022; 22(1): 59. doi: 10.1186/s12906-022-03539-8.
CrossRef - Guo Y., Zhang P., Liu Y., Zha L., Ling W., Guo H. A dose-response evaluation of purified anthocyanins on inflammatory and oxidative biomarkers and metabolic risk factors in healthy young adults: A randomized controlled trial. Nutrition. 2020; 74: 110745. doi: 10.1016/j.nut.2020.110745.
CrossRef - Hassellund S. S., Flaa A., Kjeldsen S. E., Seljeflot I., Karlsen A., Erlund I., Rostrup M. Effects of anthocyanins on cardiovascular risk factors and inflammation in pre-hypertensive men: A double-blind randomized placebo-controlled crossover study. Journal of Human Hypertension. 2013; 27(2): 100–106. doi: 10.1038/jhh.2012.4
CrossRef - Stamp L.K., Chapman P., Frampton C., Duffull S.B., Drake J., Zhang Y., Neogi T., Lack of effect of tart cherry concentrate dose on serum urate in people with gout, Rheumatology. 2020; 59(9):2374–2380, https://doi.org/10.1093/rheumatology/kez606
CrossRef - Horbowicz M., Kosson R., Grzesiuk A., Dębski H. Anthocyanins of fruits and vegetables-their occurrence, analysis and role in human nutrition. Crops Res. Bull. 2008; 68: 5-22. https://doi.org/10.2478/v10032-008-0001-8
CrossRef - McAnulty L. S. Collier S. R, Landram M. J, Whittaker D. S. Isaacs S. E. Klemka J. M., Cheek S. L., Arms J. C., McAnulty S. R. Six weeks daily ingestion of whole blueberry powder increases natural killer cell counts and reduces arterial stiffness in sedentary males and females. Nutrition Research. 2014; 34(7): 577–584. doi: 10.1016/j.nutres.2014.07.002.
CrossRef - Wrolstad R. E. Durst R. W., Lee J. Tracking color and pigment changes in anthocyanin products. Trends Food Sci. Technol.2005; 16 :423–428. doi:10.1016/j.tifs.2005.03.019
CrossRef - Wahyuningsih S., Wulandari L., Wartono M. W., Munawaroh H., Ramelan A. H. The effect of pH and color stability of anthocyanin on food colorant. In IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK. 2017; 193: 012047. doi:1088/1757-899X/193/1/012047
CrossRef - Miguel M. G. Anthocyanins: Antioxidant and/or anti-inflammatory activities. Appl. Pharm. Sci.2011; 1:7–15.
- Tena N., Martín J., Asuero A. G. State of the art of anthocyanins: Antioxidant activity, sources, bioavailability, and therapeutic effect in human health. 2020; 9: 451. doi: 10.3390/antiox9050451.
CrossRef - Kähkönen M. P., Heinonen M. Antioxidant activity of anthocyanins and their aglycons. Agric. Food Chem.2003; 51: 628–633. doi: 10.1021/jf025551i
CrossRef - Bendokas V., Stanys V., Mažeikienė I., Trumbeckaite S., Baniene R., Liobikas J. Anthocyanins: From the Field to the Antioxidants in the Body. 2020: 9 :819. doi: 10.3390/antiox9090819.
CrossRef - Ma Z., Du B., Li J., Yang Y., Zhu F. An insight into anti-inflammatory activities and inflammation related diseases of anthocyanins: A review of both in vivo and in vitro investigations. J. Mol. Sci.2021; 22: 11076. doi: 10.3390/ijms222011076.
CrossRef - Palungwachira P., Tancharoen S., Phruksaniyom C., Klungsaeng S., Srichan R., Kikuchi K., Nararatwanchai T. Antioxidant and anti-inflammatory properties of anthocyanins extracted from Oryza sativa in primary dermal fibroblasts. Oxidative Med. Cell. Longev.2019; 1 :2089817. doi: 10.1155/2019/2089817.
CrossRef - Nikbakht E., Singh I., Vider J., Williams L. T., Vugic L., Gaiz A., Kundur A. R., Colson N. Potential of anthocyanin as an anti-inflammatory agent: A human clinical trial on type 2 diabetic, diabetic at-risk and healthy adults. Res.2021; 70: 275–284. doi: 10.1007/s00011-021-01438-1.
CrossRef - Rechner A. R., Kroner C. Anthocyanins and colonic metabolites of dietary polyphenols inhibit platelet function. Thromb. Res. 2005; 116: 327-334. doi: 10.1016/j.thromres.2005.01.002.
CrossRef - Wallace T. C. Anthocyanins in cardiovascular disease. Nutr.2011; 2: 1–7. doi: 10.3945/an.110.000042.
CrossRef - Sancho R. A. S., Pastore G. M. Evaluation of the effects of anthocyanins in type 2 diabetes. Food Res. Int.2012; 46: 378–386.
CrossRef - Różańska D., Regulska-Ilow B. The significance of anthocyanins in the prevention and treatment of type 2 diabetes. Clin. Exp. Med.2018; 27:135–142. doi: 10.17219/acem/64983.
CrossRef - Nomi Y., Iwasaki-Kurashige K., Matsumoto H. Therapeutic effects of anthocyanins for vision and eye health. s2019; 24 :3311. doi: 10.3390/molecules24183311.
CrossRef - Khoo H. E., Ng H. S., Yap W S, Goh H J H, Yim H S. Nutrients for prevention of macular degeneration and eye-related diseases. Antioxidants. 2019; 8: 85. doi: 10.3390/antiox8040085.
CrossRef - Khan M. S., Ali T., Kim M. W., Jo M. H., Chung J. I., Kim M. O. Anthocyanins improve hippocampus-dependent memory function and prevent neurodegeneration via JNK/Akt/GSK3β signaling in LPS-treated adult mice. Neurobiol.2019; 56:671–687. doi: 10.1007/s12035-018-1101-1.
CrossRef - Andres-Lacueva C., Shukitt-Hale B., Galli R. L., Jauregui O., Lamuela-Raventos R. M., Joseph J. A. Anthocyanins in aged blueberry-fed rats are found centrally and may enhance memory. Neurosci.2005; 8 :111–120. doi: 10.1080/10284150500078117.
CrossRef - Alappat B., Alappat J. Anthocyanin pigments: Beyond aesthetics. 2020; 25: 5500. https://doi.org/10.3390/ molecules25235500
CrossRef - Chanoca A., Kovinich N., Burkel B., Stecha S., Bohorquez-Restrepo A,, Ueda T., Eliceiri K. W., Grotewold E., Otegui M. S. Anthocyanin vacuolar inclusions form by a microautophagy mechanism. Plant Cell 2015; 27:2545–2559. doi: 10.1105/tpc.15.00589.
CrossRef - Khoo H. E., Azlan A., Tang S. T, Lim S. M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017;61. doi: 10.1080/16546628.2017.1361779.
CrossRef - Pervaiz T., Songtao J., Faghihi F., Haider M. S., Fang J. Naturally occurring anthocyanin, structure, functions and biosynthetic pathway in fruit plants. Plant Biochem. Physiol. 2017; 5: 2. DOI: 10.4172/2329-9029.1000187
CrossRef - Kay C. Analysis of the bioactivity, metabolism, and pharmacokinetics of anthocyanins in humans. PhD thesis. University of Guelph, Ontario, Canada. 2004; 1-9.
- Muselík J., García-Alonso M., Martín-López M. P., Žemlička M., Rivas-Gonzalo J. C. Measurement of antioxidant activity of wine catechins, procyanidins, anthocyanins and pyranoanthocyanins. J. Mol. Sci. 2007; 8: 797-809. https://doi.org/10.3390/i8080797.
CrossRef - Tan Y. F., Wang M., Chen Z. Y., Wang L., Liu X. H. Inhibition of BRD4 prevents proliferation and epithelial–mesenchymal transition in renal cell carcinoma via NLRP3 inflammasomeinduced pyroptosis. Cell Death & Disease.2020; 11 (4):239. https://doi.org/10.1038/s41419-020-2431-2.
CrossRef - Krga I., Tamaian R., Mercier S., Boby C., Monfoulet L. E., Glibetic M., Morand C., Milenkovic D. Anthocyanins and their gut metabolites attenuate monocyte adhesion and transendothelial migration through nutrigenomic mechanisms regulating endothelial cell permeability. Free Radical Biology and Medicine. 2018;124: 364–379. doi: 10.1016/j.freeradbiomed.2018.06.027.
CrossRef - Molagoda I. M. N., Lee K. T., Choi Y. H., Kim G. Y. Anthocyanins from Hibiscus syriacus L. Inhibit Oxidative Stress-Mediated Apoptosis by Activating the Nrf2/HO-1 Signaling Pathway. Antioxidants, 2020;9(1). https://doi.org/10.3390/antiox9010042.
CrossRef - Taverniti V., Fracassetti D., Del Bo’ C., Lanti C., Minuzzo M., Klimis-Zacas D., Riso P., Guglielmetti S. Immunomodulatory effect of a wild blueberry anthocyanin-rich extract in human Caco-2 intestinal cells. Journal of Agricultural and Food Chemistry. 2014; 62(33) :8346–8351.
CrossRef - Liu W., Xu J., Wu S., Liu Y., Yu X., Chen J., Tang X., Wang Z., Zhu X., Li X. Selective Anti-Proliferation of HER2-Positive Breast Cancer Cells by Anthocyanins Identified by High-Throughput Screening. PLOS ONE. 2013; 8(12): e81586. doi: 10.1371/journal.pone.0081586.
CrossRef - Wongwichai T., Teeyakasem P., Pruksakorn D., Kongtawelert P., Pothacharoen P. Anthocyanins and metabolites from purple rice inhibit IL-1β-induced matrix metalloproteinases expression in human articular chondrocytes through the NF-κB and ERK/MAPK pathway. Biomedicine & Pharmacotherapy. 2019; 112 :108610. doi: 10.1016/j.biopha.2019.108610.
CrossRef - Joshi R, Rana A, Kumar V, Kumar D, Padwad Y S, Yadav S K, Gulati A. Anthocyanins enriched purple tea exhibits antioxidant, immunostimulatory and anticancer activities. Journal of Food Science and Technology. 2017; 54(7) :1953–1963. doi: 10.1007/s13197-017-2631-7
CrossRef - Zhou L, Wang H, Yi J, Yang B, Li M, He D, Yang W, Zhang Y, Ni H. Anti-tumor properties of anthocyanins from Lonicera caerulea ‘Beilei’ fruit on human hepatocellular carcinoma: In vitro and in vivo Biomedicine & Pharmacotherapy. 2018; 104: 520–529. https://doi.org/10.1016/j.biopha.2018.05.057
CrossRef - Cremonini E., Daveri, E., Mastaloudis, A., Adamo, A. M., Mills, D., Kalanetra, K., Hester, S. N., Wood, S. M., Fraga, C. G., & Oteiza, P. I. Anthocyanins protect the gastrointestinal tract from high fat diet-induced alterations in redox signaling, barrier integrity and dysbiosis. Redox Biology. 2019; 26 :101269. doi: 10.1016/j.redox.2019.101269.
CrossRef - Mazewski C., Kim, M. S., & Gonzalez de Mejia, E. Anthocyanins, delphinidin-3-O-glucoside and cyanidin-3-O-glucoside, inhibit immune checkpoints in human colorectal cancer cells in vitro and in silico. Scientific Reports. 2019; 9(1):11560. doi: 10.1038/s41598-019-47903-0.
CrossRef - Forbes-Hernández T. Y., Gasparrini M., Afrin S., Cianciosi D., González-Paramás A. M., Santos-Buelga C., Mezzetti B., Quiles J. L., Battino M., Giampieri F., Bompadre S. Strawberry (cv. Romina) Methanolic Extract and Anthocyanin-Enriched Fraction Improve Lipid Profile and Antioxidant Status in HepG2 Cells. International Journal of Molecular Sciences. 2017; 18(6). doi: 10.3390/ijms18061149.
CrossRef - Huang W., Yan Z., Li D., Ma Y., Zhou J., Sui Z. Antioxidant and Anti-Inflammatory Effects of Blueberry Anthocyanins on High Glucose-Induced Human Retinal Capillary Endothelial Cells. Oxidative Medicine and Cellular Longevity. 2018; e1862462. doi: 10.1155/2018/1862462.
CrossRef - Semmarath W., Mapoung S., Umsumarng S., Arjsri P., Srisawad K., Thippraphan P., Yodkeeree S., Dejkriengkraikul P. Cyanidin-3-O-glucoside and Peonidin-3-O-glucoside-Rich Fraction of Black Rice Germ and Bran Suppresses Inflammatory Responses from SARS-CoV-2 Spike Glycoprotein S1-Induction In Vitro in A549 Lung Cells and THP-1 Macrophages via Inhibition of the NLRP3 Inflammasome Pathway. Nutrients. 2022; 14(13). doi: 10.3390/nu14132738.
CrossRef - Ferrari D., Cimino F., Fratantonio D., Molonia M. S., Bashllari R., Busà R., Saija A., Speciale A. Cyanidin-3-O-Glucoside Modulates the In Vitro Inflammatory Crosstalk between Intestinal Epithelial and Endothelial Cells. Mediators of Inflammation. 2017(1); 3454023. doi: 10.1155/2017/3454023.
CrossRef - López de las Hazas M. C., Mosele J. I., Macià A., Ludwig I. A., Motilva M. J. Exploring the Colonic Metabolism of Grape and Strawberry Anthocyanins and Their in Vitro Apoptotic Effects in HT-29 Colon Cancer Cells. Journal of Agricultural and Food Chemistry. 2017; 65(31): 6477–6487. doi: 10.1021/acs.jafc.6b04096
CrossRef - Lo Vasco V. R., Leopizzi M., Di Mai V., Di Raimo T., Cesa S., Masci A., Rocca C. D. LPS, Oleuropein and Blueberry extracts affect the survival, morphology and Phosphoinositide signalling in stimulated human endothelial cells. Journal of Cell Communication and Signaling. 2017; 11(4) :317–327. doi: 10.1007/s12079-017-0391-9. doi: 10.1007/s12079-017-0391-9.
CrossRef - Calland N., Sahuc M. E., Belouzard S., Pène V., Bonnafous P., Mesalam A. A., Deloison G., Descamp V., Sahpaz S., Wychowski C., Lambert O., Brodin P., Duverlie G., Meuleman P., Rosenberg A. R., Dubuisson J., Rouillé Y., Séron K. Polyphenols Inhibit Hepatitis C Virus Entry by a New Mechanism of Action. Journal of Virology. 2015; 89(19), 10053–10063. doi: 10.1128/JVI.01473-15.
CrossRef - Ortiz T., Argüelles-Arias F., Begines B., García-Montes J. M., Pereira A., Victoriano M., Vázquez-Román V., Pérez Bernal J. L., Callejón R. M., De-Miguel M., Alcudia A. Native Chilean Berries Preservation and In Vitro Studies of a Polyphenol Highly Antioxidant Extract from Maqui as a Potential Agent against Inflammatory Diseases. Antioxidants. 2021; 10(6). doi: 10.3390/antiox10060843.
CrossRef - Amini A. M., Muzs K., Spencer J. P. E., Yaqoob P. Pelargonidin-3-O-glucoside and its metabolites have modest anti-inflammatory effects in human whole blood cultures. Nutrition Research. 2017; 46:88–95. doi: 10.1016/j.nutres.2017.09.006.
CrossRef - Guo Y., Zhang P., Liu Y., Zha L., Ling W., Guo H. A dose-response evaluation of purified anthocyanins on inflammatory and oxidative biomarkers and metabolic risk factors in healthy young adults: A randomized controlled trial. Nutrition. 2020;74 :110745. doi: 10.1016/j.nut.2020.110745.
CrossRef - Cremonini E., Daveri E., Iglesias D. E., Kang J., Wang Z., Gray R., Mastaloudis A., Kay C. D., Hester S. N., Wood S M, Fraga C G, Oteiza P I. A randomized placebo-controlled cross-over study on the effects of anthocyanins on inflammatory and metabolic responses to a high-fat meal in healthy subjects. Redox Biology. 2022; 51: 102273. doi: 10.1016/j.redox.2022.102273.
CrossRef - Zhang H., Xu Z., Zhao H., Wang X., Pang J., Li Q., Yang Y., Ling W. Anthocyanin supplementation improves anti-oxidative and anti-inflammatory capacity in a dose–response manner in subjects with dyslipidemia. Redox Biology. 2020; 32: 101474. doi: 10.1016/j.redox.2020.101474.
CrossRef - do Rosario V. A., Chang C., Spencer J., Alahakone T., Roodenrys S., Francois M., Weston-Green K., Hölzel N., Nichols D. S., Kent K., Williams D., Wright I. M. R., Charlton K. Anthocyanins attenuate vascular and inflammatory responses to a high fat high energy meal challenge in overweight older adults: A cross-over, randomized, double-blind clinical trial. Clinical Nutrition. 2021a; 40(3):879–889. doi: 10.1016/j.clnu.2020.09.041.
CrossRef - Curtis P.J., Kroon P.A., Hollands W.J., Walls R., Jenkins G., Kay C.D., Cassidy A. Cardiovascular disease risk biomarkers and liver and kidney function are not altered in postmenopausal women after ingesting an elderberry extract rich in anthocyanins for 12 weeks. J Nutr. 2009;139(12):2266-71. doi: 10.3945/jn.109.113126.
CrossRef - Aboonabi A., Meyer R. R., Gaiz A., Singh I. Anthocyanins in berries exhibited anti-atherogenicity and antiplatelet activities in a metabolic syndrome population. Nutrition Research.2020; 76: 82–93. doi: 10.1016/j.nutres.2020.02.011.
CrossRef - Willems M. E., Myers S. D., Gault M. L., Cook M. D. Beneficial Physiological Effects With Blackcurrant Intake in Endurance Athletes. Int J Sport Nutr Exerc Metab. 2015 ;25(4):367-74. doi: 10.1123/ijsnem.2014-0233.
CrossRef - Pekas E. J., Shin J., Headid R. J., Son W. M., Layec G., Yadav S. K., Scott S. D., Park S.Y. Combined anthocyanins and bromelain supplement improves endothelial function and skeletal muscle oxygenation status in adults: A double-blind placebo-controlled randomised crossover clinical trial. British Journal of Nutrition. 2021; 125(2): 161–171. doi: 10.1017/S0007114520002548.
CrossRef - Hassellund S. S., Flaa A, Kjeldsen S E, Seljeflot I, Karlsen A, Erlund I, Rostrup M. Effects of anthocyanins on cardiovascular risk factors and inflammation in pre-hypertensive men: A double-blind randomized placebo-controlled crossover study. Journal of Human Hypertension. 2013; 27(2): 100–106. doi: 10.1038/jhh.2012.4.
CrossRef - do Rosario V. A., Fitzgerald Z., Broyd S., Paterson A., Roodenrys S., Thomas S., Bliokas V., Potter J., Walton K., Weston–Green K., Yousefi M., Williams D., Wright I. M. R., Charlton K. Food anthocyanins decrease concentrations of TNF-α in older adults with mild cognitive impairment: A randomized, controlled, double blind clinical trial. Nutrition, Metabolism and Cardiovascular Diseases. 2021b; 31(3): 950–960. doi: 10.1016/j.numecd.2020.11.024.
CrossRef - Şahin M. A., Bilgiç P., Montanari S., Willems M.E.T. Intake Duration of Anthocyanin-Rich New Zealand Blackcurrant Extract Affects Metabolic Responses during Moderate Intensity Walking Exercise in Adult Males. Journal of Dietary Supplements. 2021; 18(4):406–417. doi: 10.1080/19390211.2020.1783421.
CrossRef - Santamarina A. B., Jamar G., Mennitti L. V., Cesar H. de C., Vasconcelos J. R., Oyama L. M., de Rosso V., Pisani L. P. Obesity-related inflammatory modulation by juçara berry (Euterpe edulis Mart.) supplementation in Brazilian adults: A double-blind randomized controlled trial. European Journal of Nutrition. 2020; 59(4): 1693–1705. doi: 10.1007/s00394-019-02024-2.
CrossRef - Nikbakht E., Singh I., Vider J., Williams L. T., Vugic L., Gaiz A., Kundur A. R., Colson N. Potential of anthocyanin as an anti-inflammatory agent: A human clinical trial on type 2 diabetic, diabetic at-risk and healthy adults. Inflammation Research.2021; 70(3):275–284. doi: 10.1007/s00011-021-01438-1.
CrossRef - McAnulty L. S., Collier S. R., Landram M. J., Whittaker D. S., Isaacs S. E., Klemka J. M., Cheek S. L., Arms J. C., McAnulty S. R. Six weeks daily ingestion of whole blueberry powder increases natural killer cell counts and reduces arterial stiffness in sedentary males and females. Nutrition Research. 2014; 34(7): 577–584. doi: 10.1016/j.nutres.2014.07.002.
CrossRef - Emamat H., Zahedmehr A., Asadian S., Nasrollahzadeh J. The effect of barberry (Berberis integerrima) on lipid profile and systemic inflammation in subjects with cardiovascular risk factors: A randomized controlled trial. BMC Complementary Medicine and Therapies. 2022; 22(1): 59. doi: 10.1186/s12906-022-03539-8.
CrossRef - Nathan C, Ding A. Nonresolving inflammation. Cell. 2010; 140 :871–882. doi: 10.1016/j.cell.2010.02.029.
CrossRef - Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008; 454: 428–435. https://doi.org/10.1038/nature07201.
CrossRef - Brown J C, Winters-Stone K, Lee A, Schmitz K H. Cancer, physical activity, and exercise. Physiol. 2012;2: 2775. doi: 10.1002/cphy.c120005.
CrossRef - Muñoz N, Castellsagué X, de González A B, Gissmann L. HPV in the etiology of human cancer. 2006: 24; S1–S10. doi: 10.1016/j.vaccine.2006.05.115.
CrossRef
Abbreviations
PRISMA- Preferred Reporting Items for Systematic reviews and Meta-Analyses, IL-1- Interleukin-1, IL- 6- Interleukin-6, IL- 10- Interleukin-10, HDL- High density lipoproteins, mRNA-Messenger ribonucleic acid, Cy3Gal – Cyanidin-3-galactoside, C3G -Cyanidin-3-glucoside, D3G- Delphinidin-3O-glucoside, P3G- Peonidin-3-glucoside, NF-κB-Nuclear factor-kappa B, TNFα- Tumor necrosis factor alpha, HaCat- High sensitivity of human epidermal keratinocytes, H2O2– Hydrogen Peroxide, ROS- Reactive Oxygen Species, Nrf2- Nuclear factor erythroid 2-related factor 2, HFD- High fat diet, NOX- Nicotinamide adenine dinucleotide phosphate (NADPH)-oxidase, ERK- extracellular signal-regulated kinase, PD-L1- Programmed Cell Death Ligand 1, HCT- Hematocrit, LDL- Low density lipoproteins, HepG2-Hepatoblastoma, SOD – Superoxide dismutase, VEGF – Vascular endothelial growth factor, SP- SARS-CoV-2 , NLRP3- Nucleotide-binding domain, leucine-rich–containing family, pyrin domain–containing-3, ASC- apoptosis-associated speck-like protein, IL-8- Interleukin-8, EC- Endothelial cell, LPS- Lipopolysaccharide, 8-OHdG-8-hydroxydeoxyguanine , 8-iso-PGF2α-8-iso-prostaglandin F2α, MDA- Malondialdehyde, BMI- Body Mass Index, CRP- C-reactive protein, CVD-Cardio vascular disease, FBG- Fasting blood sugar, TG-Triglyceride, MCP-1 – Monocyte chemoattractant protein-1 , IL-10 – Interleukin-10, NK- Natural killer cells , Aix- augmentation index, ASP- aortic systolic pressures, T2DM- Type 2 diabetes mellitus.

This work is licensed under a Creative Commons Attribution 4.0 International License.






