Effect of in Vitro Gastrointestinal Digestion on Bioactivity of Poultry Protein Hydrolysate
1Federal Research Centre “Fundamentals of Biotechnology” of The Russian Academy of Sciences”, 119071, Russia, Moscow, Leninskiy Prospekt, 33, bld. 2 2Moscow Institute of Physics and Technology, Moscow, Russia; Dolgoprudnyi, 141700 Moscow Region, Russia 3Emanuel Institute for Biochemical Physics, Russian Academy of Sciences, 119334, Kosygina street, 4, Moscow, Russia 4Institute of General Pathology and Pathophysiology, 125315, Baltiysaya street, 8, Moscow, Russia.
Corresponding Author Email: snowsurfers1@gmail.com
DOI : http://dx.doi.org/10.12944/CRNFSJ.4.Special-Issue-October.10
Download this article as:
![]()
In vitro simulated gastrointestinal digestion (GID) was performed to evaluate changes in bioactive properties of Poultry protein hydrolysate HCP Premium P150 (PPH) showing strong antioxidant (448.2±37.0 µM TE/g of protein) and moderate Angiotensin-I converting enzyme inhibitory activity (IC50 0.617±0.022 mg/ml). Antioxidant and ACE-inhibitory activity were measured with use of ORAC assay and FRET-substrate methods, correspondingly. Gastric digestion (GD) increased ACE inhibitory activity 2.23 times and didn’t change antioxidant activity of PPH significantly. The subsequent intestinal digestion increased antioxidant activity 1.29 times and didn’t change ACE-inhibitory activity significantly. New potent ACE-inhibitory peptides: APGAPGPVG (IC50 16.2±3.8 µM), PDLVF (IC50 84.9±6.3 µM) and antioxidant dipeptide WG (2.29±0.04 µM TE/µM) were identified in the digested PPH. The digested PPH proved to be a rich source of antioxidant and ACE inhibiting molecules and could be a potential new food ingredient used for prevention or treatment of socially significant diseases.
KEYWORDS:Antioxidant hydrolysate; ACE-inhibitory hydrolysate; in vitro gastrointestinal digestion; chicken hydrolysate
Introduction
The growth of common diseases such as cardiovascular disorders, strokes, diabetes, cancer and atherosclerosis has become a serious global problem. In the last decade, the researches paid considerable attention to the use of certain food components, especially bioactive peptides, to prevent socially significant diseases or reduce their negative effects. The use of biopeptides and bioactive hydrolysates as functional food ingredients is a promising solution for some health problems. In contrast with synthetic drugs, peptides display very few side effects in humans due to their natural origin. Bioactive peptides may be released during in vivo digestion or food processing steps, i.e. fermentation, ripening, in vitro enzymatic hydrolysis1. Reсently, dairy proteins have been the most studied source of bioactive peptides2. However, other proteins of animal origin, such as collagens of skins, muscle myosins, and sarcoplasmic proteins have also been shown to have varying potential biological effects.
Chicken is a significant source of food and plays an important role in the economy of many countries. Its processing industry produces huge amounts of waste which are usually discarded or rarely processed into hydrolysates and used as food-producing animal protein supplements. Such hydrolysates are perspective sources of molecules with bioactive properties such as ACE-inhibitory or antioxidant peptides and may be used for prevention or treatment of cardiovascular or cancer diseases, organism aging, and degenerative diseases3. The hydrolysate of Gallus Gallus collagen and muscle proteins, HCP P150 (PPH), was previously designed in the course of work on poultry meat-and-bone residues bioconversion with the use of an enzyme composition containing Alcalase, Protamex, Neutrase, and Flavourzyme4. The PPH has shown strong antioxidant effect in vitro.
After oral administration, during passage through the gastro-intestinal tract, peptide profiles of hydrolysates and consequently their bioactive properties may suffer serious changes before reaching the target organs. Resistance of peptides to proteolysis by gastrointestinal, brush border and serum peptidases is important. In our previous studies5, Nikolaev et al. observed a mild antioxidant effect of PPH in the central nervous system in vivo, in Wistar-Kyoto rats. However, the bioactive peptides profile in PPH remains unclear, as well as the peptide degradation processes during the passage through the digestive tract.
The objective of this study was to evaluate the stability of antioxidant and ACE-inhibitory properties of hydrolysate HCP Premium 150 (PPH) (Proliver, Belgium) when exposed to digestive enzymes, and on the base of amino acid descriptors of antioxidant and ACE inhibitory activities to analyze the hydrolysate components responsible for bioactivity.
Materials and methods
Poultry hydrolysate HCP Premium (PPH) ( Proliver, Belgium);
Trypsin from hog pancreas, 11909 U/Mg (93614 Fluka BioChemika);
Pepsin from porcine gastric mucosa, 3200-4500 units/mg protein (P 6887 Sigma);
α-Chymotrypsin from bovine pancreas, 60 U/mg protein (Sigma C 4129);
Abz-FRK(Dnp)-OH (A4980 Sigma);
Abz-LFK(Dnp)-OH (A5855 Sigma);
Peptides H-His-Tyr-Val-Pro-Val-OH, H-Pro-Asp-Leu-Val-Phe-OH, H-Ala-Pro-Gly-Ala-Pro-Gly-Pro-Val-Gly-OH, H-Trp-Gly-OH, H-Met-Lys-OH, and H-Met-Met-OH with purity >98% were purchased from Bachem Budendorf, Switzerland.
Evaluation of in vitro hypotensive activity
ACE-inhibitory activity was measured according to the method of Carmona et al.6 with use of Abz-FRK(Dnp)P-OH and Abz-LFK(Dnp)P as FRET substrates for hydrolysates and peptides, correspondingly. ACE from rabbit lung (Sigma) was mixed with serial dilutions of hydrolysates or peptide solutions in wells of 96-well black microplate (Greiner bio) with a final ACE concentration of 0.01 U/ml. Microplate was incubated and stirred for 30 min at 37°C. After incubation, FRET substrates in concentration 10µM were added to each well. Total volume of reaction solution was 200µL. Fluorescence intensity was monitored using Synergy 2 (BioTek) spectrophotometer-fluorimeter for 30 min at the following working parameters of the reader: excitation at 320 nm, emission filter at 420 nm, reading interval 45s, temperature 37 °C, and constant stirring. ACE residual activity was calculated as follows:
ACE residual activity (%) = Ainhibitor x 100/Acontrol
The half maximal inhibitory concentration (IC50) was used to express the potency of the sample to inhibit ACE activity. The IC50 value was expressed in mg of protein/ml for hydrolysates and in µM for peptides.
Determination of antioxidant capacity of the hydrolyzates by ORAC-FL assay
ORAC assay was based on the method of Moor et al.7 modified by Koroleva et al.8 Trolox solutions (5–75 μM) were used for calibration. Fluorescence intensity was monitored for 1 hour under the following working parameters of the reader: excitation at 485 nm, emission filter at 528 nm, reading interval 60 s. The final ORAC-FL values of hydrolysates were calculated by using the linear regression equation between the trolox concentration and the net area (net AUC) of the fluorescein decay curve (net AUC=Rf [trolox]). The ORAC values were expressed in µMTE/g for the hydrolysates and in µMTE/ µmol for peptides.
Simulated gastric juice
The simulated gastric acid solution was prepared on the base of USP 26 standard and contained HCl (0.23 M) and NaCl (0.034 M). The simulated gastric enzyme solution was prepared by dissolving porcine gastric mucosa pepsin (activity 3411 U/mg of protein calculated using hemoglobin as substrate, Sigma) in the described salt mixture to reach the final concentration of 8000U/mL pepsin.
Simulated pancreatic juice
The simulated pancreatic juice was prepared on the base of USP 26 and Pitino et al9. Pancreatic juice solution contained NaCl (0.15 M) and KH2PO4 (0.05 M). Trypsin from hog pancreas (activity 11,909 U/mg of protein calculated using BAEE as substrate, Fluka BioChemika), a-chymotrypsin from bovine pancrease (activity 40 U/mg of protein calculated using BTEE as substrate, Novozymes, Belgium) were added to pancreatic juice so that the final enzyme concentrations were: 11 U/mL for Hog trypsin and 24 U/mL for bovine a-chymotrypsin.
Simulated gastrointestinal digestion
Chicken hydrolysate was dissolved in the gastric juice to reach protein concentration of 20 mg/ml. pH was adjusted to 1.8. The mixture was incubated at 37°C for 1 hour with constant stirring. Upon completion of the hydrolysis, the pH of the solution was adjusted to 6.8 pH to inactivate the enzyme.
20 ml of the neutralized mixture after gastric hydrolysis was quantitatively transferred into a flask containing 10 ml of intestinal juice. pH was adjusted to 6.8 and solutions of trypsin and chymotrypsin were added and incubated at 37 °C for 2 hours. The inactivation of enzyme preparations after intestinal hydrolysis was carried out in a water bath at 80 °C for 15 minutes.
Determination of molecular mass distribution
Molecular mass distribution was analyzed using a BioSep S 2000 column (7.8×300 mm) from Phenomenex (Torrance, Ca, USA) installed into the ProStar HPLC chromatographic device (Varian Inc., USA). Protein and peptide kits from GE Healthcare (USA), Sigma (USA) and Serva (Germany) were used as standards for calibration. The hydrolysates were filtered through 0.45 µm hydrophilic filters (Sartorius AG, Germany), and were injected (20 µL) into the system with subsequent elution with 50 mM potassium phosphate buffer, pH 6.9, at a flow rate of 1 ml/min. The UV-Vis detector was set at 214 nm.
Peptide profile analysis
Peptide profile analysis was performed by means of high performance reversed-phase chromatography with electrospray ionization. The components were identified by mass spectrometry of ion cyclotron resonance with Fourier transform. Fractions were sub-fractioned in a liquid chromatographic system Agilent 1100 (Agilent Technogies, Paolo Alto, CA, USA) on a capillary column (inner diameter 75 µm x length 12 cm) containing Reprosil-Pur Basic C18 media, 3 µm (Dr. Maisch HPLC GmbH, Ammerbuch-Entringen, Germany).
The detectors of the chromatographic system consisted of a mass spectrometer 7-Tesla Finnigan LTQ-FT Ultra (Thermo Electron, Germany) equipped with a nanoelectrospray. The Xcalibur software package (Thermo Finnigan, USA) was used for experimental data registration.
The Mascot Daemon 2.2.2 (Matrix Science, UK) software was used for automatic searching of the UniProtKB_SwissProt sprot_v.57.9 database. Acceptable level of significance was set at 95% probability. The peptides were considered reliably identified if Mascot score was at least 24.
Statistic analysis
The measurement of antioxidant capacity and ACE inhibition concentration values was performed in 4 replicates. The results are given as mean ± sd.
During the analysis of MW distribution in the hydrolysate fractions, the optical density was measured in 4 replicates. The content of fractions is given as mean values.
Results and discussion
Changes of MW distribution after simulated digestion of PPH
It was previously shown that the hydrolysate of collagen and muscle proteins, HCP Premium 150 (PPH), had a pronounced antioxidant activity in vitro4,5 and a moderate antioxidant effect in vivo5. However, the bioactive peptides profile in PPH remains unclear, as well as the peptide degradation processes during the passage through the digestive tract.
An in vitro model was designed on the basis of USP 26 protocol to assess the changes of hydrolysate bioactivity properties during digestion by the enzymes of the gastrointestinal tract (GIT). The in vitro model included consecutive processing of PPH first by artificial gastric (GD), and then by intestinal juice (GID), to obtain experimental digestive protein hydrolysates, DPHexp-GD and DPHexp-GID. Molecular weight distribution analysis was performed for PPH, DPHexp-GD, and DPHexp-GID. After GID, the PPH fraction containing peptides >3 kDa was reduced from 45.54% to 36.03% (Table 1). The HPLC profiles of PPH before and after the GID are similar, but the digestion results in a decrease of the peak intensity with MW of 3800 Da, while the intensity of the peak with MW 1500 Da increases (Fig 1). Also a new peak with MW 170 Da, corresponding to the MW of di – and tripeptides appears in the DPHexp-GID sample. Specificity of trypsin and chymotrypsin is wider than that of pepsin10,11, which allows to obtain fractions with a lower MW.
Table 1: Molecular weight distribution of the hydrolysates.
| Sample | Content of the fraction, % | ||
| <3 kDa | 3-10 kDa | >10 kDa | |
| PPH | 54.46 | 41.20 | 4.34 |
| DPHexp-GD | 59.96 | 35.13 | 4.91 |
| DPHexp-GID | 63.97 | 34.19 | 1.84 |
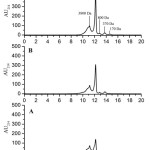 |
Figure 1: HPLC chromatogram of PPH (A), DPHexp-GD (B), DPH exp-GID (C). Click here to View figure |
In vitro analysis of PPH bioactivity after processing by gastrointestinal enzymes
Comparison of bioactive properties in PPH, DPHexp-GD, and DPHexp-GID showed a statically significant (P<0.05) increase of ACE inhibitory activity (IC50 of PPH 0.617±0.022 mg/ml) after gastric digestion (IC50 of DPHexp-GD 0.270±0.016 mg/ml), while antioxidant activity was not influenced (Fig 2). Subsequent intestinal digestion (GID) increased antioxidant activity 1.29 times (P<0.05), from 486.5±25.8 µM TE/g of protein for DPHexp-GD to 628.1±32.8 µM TE/g of protein for DPHexp-GID (Fig 2). After GID, the ACE-inhibitory activity didn’t change significantly compared to GD. This is probably due to the appearance of a peak with MW 170 Da, corresponding to the di- and tripeptides. It appears that dipeptides which are more abundant in DPHexp-GID (MW 170 Da) form weaker contacts with the active site of the enzyme, while tripeptides and longer molecules inhibit ACE more effectively12. Meanwhile, a dipeptide sequence is probably sufficient for effective quenching of free radical reactions that may cause an increase in AOE after GID (Fig 2).
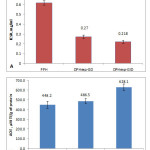 |
Figure 2: Bioactivity of PPH and its digested forms, DPHexp-GD and DPHexp-GID: A) ACE-inhibitory activity, B) antioxidant activity. Click here to View figure |
Similar results were observed in report of Senphan and Benjakul13 when ABTS radical scavenging activity and chelating activity of sea bass skin hydrolysate was slightly increased during pepsin digestion, but extremely increased within the first 20 min under duodenal conditions. The ACE inhibitory properties of the sardine and tuna by-products hydrolysates were greatly improved by in vitro gastric digestion, and were barely affected by further intestinal digestion14, which is in good agreement with results observed in our work. However, Ketnawa et al.15 found that the ABTS radical scavenging activity observed after GID Gelatin hydrolysates from farmed Giant Catfish skin varied significantly depending on the enzyme used for gelatin hydrolysis, but generally increased. The antioxidant activity of porcine trypsin hydrolysate mainly increased during the gastric digestion. In contrast, the ABTS radical scavenging activity of Izyme AL® hydrolysate slightly increased during gastric digestion and significantly increased during the intestinal digestion15. Espejo-Carpio et al.16 reported that intestinal digestion altered the peptide profiles and bioactivities of goat milk hydrolysates, whereas the effect of the gastric digestion was negligible. They observed a decrease of ACE inhibitory activity after GID when peptides were produced with trypsin, and an increase of antioxidant activity when either trypsin or subtilisin were used for raw material hydrolysis16. It indicates that effects of GID on bioactivity are associated not only with specificity of digestive enzymes and MW decrease of peptides17,18, but also with sequences and conformation of peptides in hydrolysates, which is related with the specificity of enzymes used in raw material hydrolysis.
Peptide profile of hydrolysates
The peptide profile of the PPH and DPHexp-GID hydrolysates was established by high performance reversed-phase chromatography with electrospray ionization. Identification of the components was performed by mass spectrometry of ion cyclotron resonance with Fourier transform. The number of identified peptides was 689 and 450 for PPH and DPHexp-GID, respectively. The minimum peptide size that can be identified by the ESI-MS-MS method is 5 amino acid residues, while peptides with high bioactivity potential with a size of 2-4 amino acid residues are not taken into account. To model the di-, tri-, and tetrapeptide formation during gastro-intestinal hydrolysis of PPH, an in silico digestion of PPH was performed using ExPASy database19, taking into account the specificity of pepsin, trypsin and chymotrypsin. The resulting hydrolysate, DPHmod, was a mixture of 722 peptides with length of 2-19 amino acid residues. The comparison of the peptide profiles showed that after in vitro simulated digestion, the peptides of sarcoplasmic origin prevailed in DPHexp-GID, while in the original PPH and its in silico digested form (DPHmod) the peptides of myofibrillar proteins prevailed. The difference is probably due to the identification limits of ESI-MS-MS method. In DPHexp-GID, 33 peptides were similar to those identified in the original PPH: 9 formed during digestion from longer peptides, 13 peptides remained undigested, and 11 peptides might form either way. 30 peptides found in DPHmod were identical to those in the original PPH and 7 peptides identical to those in DPHexp-GID.
In silico analysis of PPH, DPHexp-GID, and DPHmod peptide profiles showed the presence of highly active ACE-inhibitory peptides WVPSV (IC50 0.37 µM), VVYPW(IC50 0.36 µM) in DPHexp-GID and less active peptides DMIPAQK (IC50 45.0 µM) and EKSYELP (IC50 14.41 µM) in PPH. Also, an antioxidant peptide WDDMEK was identified in both DPHexp-GID and PPH. Only the dipeptides20 with biological activity were predicted in DPHmod(Table2).
Table 2: Bioactive peptides with previously reported properties identified in DPHexp-GID and predicted in DPHmod.
| Hydrolysate | ACE-inhibitory peptides | Antioxidant peptides |
| PPH | DMIPAQK, EKSYELP21, 22 | WDDMEK33 |
| DPHexp-GID | WVPSV, VVYPW23, 24 | |
| DPHmod | AY, AI, PQ, GY, LF, VG, IG, GA, GL, GH, GR, GK, GT, GE, SG, GD, VR, QK, NY, SY, KY, AR, EI, IE, EV, VE, TQ, AH, EK, WG25-32 | AY, AH34, 35 |
New bioactive peptides
It seems likely that aside from a few peptides with annotated biological activity, high antioxidant and ACE-inhibitory activity of the digested PPH are determined by a number of other peptides with previously unstudied properties.
The amino acid side chains mostly determine the bioactivity of the peptides. Sagardia et al. established a QSAR model for oligopeptides and showed that the most important factors for ACE inhibitory activity are: the presence of aliphatic bulky side chain amino acid residues such as I, L, V, A, G for C1 terminal position; T, R, V for C2; polar and charged amino acids as D, N, K, and not hydrophobic amino acids L, M, F for C3; W, Y, C, M for C4; aliphatic amino acids V, I, A for C5 position36. Also their work36 has shown that C1 and C4 terminal position are the most important positions for ACE inhibition potency, which is in agreement with an earlier work of Wu et al37. In a number of papers the preference of hydrophobic amino acids I, L, V and aromatic amino acids W, F, Y was established for ACE-inhibitory activity of di- and tripeptides38-41. It has also been reported that P at C1 and C2 positions is important both for ACE-inhibition and for resisting the proteolysis by gastrointestinal enzymes and serum peptidases11,12,39,41. Many agree that the presence of highly hydrophobic amino acids and especially aromatic amino acids in peptides, such as W, Y, M, L, T, and F, have a great importance for antioxidant properties of di- and tripeptides as well as longer peptides, especially for C3 position in the case of peroxyl radical scavenging activity (ORAC)5,42,43. The electrons in the resulting radicals are delocalized by the peptide bond or functional groups of the side chain, i.e. tryptophan aromatic ring, making these radicals more stable and less reactive44.
70 peptides with high biological activity potential were selected based on the reported descriptors, of which 6 were synthesized for subsequent verification of the effects in vitro. The ability of each peptide to inhibit the C-domain of ACE, responsible for the regulation of blood pressure, was evaluated, as well as its antioxidant activity against the peroxyl radical. Characteristics of these peptides are shown in table 3.
Table 3: Characteristics of the synthesized bioactive peptides identified in DPHexp-GID and predicted in DPHmod.
| Peptide | IC50 for C-domain of ACE, µM | Antioxidant activity (ORAC) µM TE/µM | Precursor protein | Precursor hydrolysate |
| APGAPGPVG | 16.2 ± 3.8 | 0.000759 ± 0.00001 | Collagen alpha-1(I) chain | DPHexp-GID |
| HYVPV | 998.2 ± 125.2 | 0.36 ± 0.09 | Collagen alpha-3(VI) chain | DPHexp-GID |
| PDLVF | 84.9 ± 6.3 | 0.0023 ± 0.0003 | Cytochrome P450 26A1 | DPHexp-GID |
| WG | 729.4 ± 45.6 | 2.29 ± 0.04 | Hemoglobin subunit alpha-β | DPHmod |
| MK | 6498.6 ± 626.0 | 0.30 ± 0.01 | α- Actin | DPHmod |
| MM | 547.5 ± 51.3 | 0.64 ± 0.03 | α- Actin | DPHmod |
APGAPGPVG peptide (IC50 16.2±3.8 µM) of collagen origin, characterized by a high content of P and the presence of V in the C-2 position, displayed the greatest ACE inhibitory activity comparable with the well-known ACE inhibitory peptides, IPP (IC50 5 µM) and VPP (IC50 9 µM), as well as other peptides of collagen origin: IAPG (IC50 11.4 µM), GPV (IC50 4.7 µM)45-47. PDLVF peptide (IC50 84.9±6.3 µM) also containing V in the C-2 position and P in N-1 position, showed a moderate ability to inhibit ACE.
HYVPV peptide (IC50 998.2±125.2 µM) showed a very weak ability to inhibit ACE, as well as dipeptides, but had the highest peroxyl radical scavenging activity among the oligopeptides, probably due to the presence of H and Y residues5. None of the selected dipeptides expressed ACE-inhibitory activity, which is probably related to the structural features of the active site of the enzyme and the mechanism of the peptide bond hydrolysis by an ACE molecule12,19.
The highest antioxidant activity was observed for the WG dipeptide (AOC 2.29±0.04 µM TE/µM), which is associated with the presence of W – the most redox-active amino acid after cystein48. Dipeptides containing M also showed medium antioxidant activity against peroxyl radical: 0.30±0.01 µM TE/µM and 0.64±0.03 µM TE/µM for MK and MM, respectively. Antioxidant activity of APGAPGPVG and PDLVF was very weak.
Conclusion
The simulated gastrointestinal digestion of the poultry protein hydrolysate PPH improved its bioactivity significantly. Analysis of the peptide profiles of in vitro and in silico gastro-intestinal digests, DPHexp-GID and DPHmod, revealed the presence of previously reported and newly discovered peptides with high ACE-inhibitory (APGAPGPVG with IC50 16.2±3.8 µM and PDLVF with IC50 84.9±6.3 µM) and antioxidant activity (WG, 2.29±0.04 µM TE/µM). Results of antioxidant capacity analysis of DPHexp-GID in vitro is in good agreement with the mild antioxidant effect in central nervous system previously discovered for Wistar rats with feed rations containing different protein ingredients (casein, PPH and protein-free diet)5. Both direct radical scavenging and indirect mechanisms (i.e. activation of intrinsic antioxidant defense system) could be involved in the observed antioxidant effect of PPH in vivo. Since the antihypertensive effects observed in vitro often disagree with in vivo tests, the ACE-inhibitory effects of hydrolysates and identified peptides should be further verified in vivo. Generally, PPH has great perspectives as a functional supplement in food industry.
Acknowledgments
We thank Proliver (Belgium) for providing the chicken hydrolysate. The study was financially supported by Grant of Russian Foundation for Basic Research № 16-33-01128. LC-MSMS analysis was performed at Core Facility of the Emanuel Institute for Biochemical Physics, Russian Academy of Sciences “New Material and Technologies.”
References
- Zou T-B, He T-P, Li H-B, Tang H-W, Xia E-Q . The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules; 21(1): 72: (2016).
CrossRef - Santiago-López L, Hernández-Mendoza A, Belinda V-C, Mata-Haro V and González-Córdova A. F Food-derived immunomodulatory peptides. Journal of the Science of Food and Agriculture; 96: 3631–3641: (2016).
CrossRef - Ryan J. T, Ross J. T, Bolton D, Fitzgerald G. F, Stanton C. Bioactive Peptides from Muscle Sources: Meat and Fish. Nutrient; 3(9): 765-791: (2011).
CrossRef - Nikolaev I. V, Sforza S, Lambertini F, Ismailova D. Y, Khotchenkov V. P, Volik V. G, Dossena A, Popov V. O, Koroleva O. V. Biocatalytic conversion of poultry processing leftovers: Optimization of hydrolytic conditions and peptide hydrolysate characterization. Food Chemystry; 197(Pt A): 611-21: (2016).
- Nikolaev I. V, Kononikhin A. S, Torkova A. A, Sforza S,. Koroleva O.V. Chapter The Strategy for Screening of Antioxidant Constituents in Protein Hydrolysates. In: Emerging and Traditional Technologies for Safe, Healthy and Quality Food; 145-160: 17 December (2015).
- Carmona A.K, Schwager S.L, Juliano M. A, Juliano L, Sturrock E.D. A continuous fluorescence resonance energy transfer angiotensin I-converting enzyme assay. Nature Protocols; 1(4): 1971-6: (2006).
CrossRef - Moore J, Cheng Z, Su L, Yu L. Effects of solid-state enzymatic treatments on the antioxidant properties of wheat bran. Journal of Agricultural and Food Chemistry; 54: 9032-9045: (2006).
CrossRef - Koroleva O. V, Torkova A, Nikolaev I, Khrameeva E, Fedorova T, Tsentalovich M, Amarowicz R. Evaluation of the Antiradical Properties of Phenolic Acids. International Journal of Molecular Sciences; 15(9): 16351-16380: (2014).
CrossRef - Pitino I, Randazzo C. L, Mandalari G, Curto A. L, Faulks R. M, Le Marc Y, Bisignano C, Caggia C, Wickham M. S. J. Survival of Lactobacillus rhamnosus strains in the upper gastrointestinal tract. Food Microbiology; 27 (8): 1121-1127: (2010).
CrossRef - Keil B. Proteolysis Data Bank: specificity of alpha-chymotrypsin from computation of protein cleavages. Protein Sequence Data Analysis; 1: 13-21: (1987).
- Keil B. Specificity of proteolysis. Springer-Verlag Berlin-Heidelberg-NewYork: 335: (1992).
- Natesh R, Schwager S.L, Sturrock E.D, Acharya K.R. Crystal structure of the human angiotensin-converting enzyme-lisinopril complex. Nature; 421(6922): 551-4: (2003).
CrossRef - Senphan T, Benjakul S. Antioxidative activities of hydrolysates from sea bass skin prepared using protease from hepatopancreas of Pacific white shrimp. Journal of Functional Foods; 6: 147–156: (2014).
CrossRef - Martínez-Alvarez O, Batista I, Ramos C, Montero P. Enhancement of ACE and prolyl oligopeptidase inhibitory potency of protein hydrolysates from sardine and tuna by-products by simulated gastrointestinal digestion. Food and Function; 7(4): 2066-73: (2016).
CrossRef - Ketnawa S, Martínez-Alvarez O, Benjakul S, Rawdkuen S. Gelatin hydrolysates from farmed Giant catfish skin using alkaline proteases and its antioxidative function of simulated gastro-intestinal digestion. Food Chemistry; 192: 34–42: 2016.
CrossRef - Espejo-Carpio F. J, García-Moreno P. J, Pérez-Gálvez R, Morales-Medina R, Guadix A, Guadix E. M. Effect of digestive enzymes on the bioactive properties of goat milk protein hydrolysates. International Dairy Journal; 54:21-28: (2016).
CrossRef - Zhang M, Mu T-H. Optimisation of antioxidant hydrolysate production from sweet potato protein and effect of in vitro gastrointestinal digestion. International Journal of Food Science and Technology; V 51(8): 1844–1850: (2016).
CrossRef - Zhang M, Mu T.H, Sun M.J. Sweet potato protein hydrolysates: antioxidant activity and protective effects on oxidative DNA damage. International Journal of Food Science and Technology; 47: 2304–2310: (2012).
CrossRef - Artimo P, Jonnalagedda M, Arnold K, Baratin D, Csardi G, de Castro E, Duvaud S, Flegel V, Fortier A, Gasteiger E, Grosdidier A, Hernandez C, Ioannidis V, Kuznetsov D, Liechti R, Moretti S, Mostaguir K, Redaschi N, Rossier G, Xenarios I, and Stockinger H. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Research; 40(1): 597-603: (2012).
CrossRef - Minkiewicz P, Dziuba J, Iwaniak A, Dziuba M, Darewicz M. BIOPEP database and other programs for processing bioactive peptide sequences. Journal of AOAC International; 91: 965-980: (2008).
- Yokoyama K, Chiba H, Yoshikawa M. Peptide inhibitors for angiotensin I-converting enzyme from thermolysin digest of dried bonito. Bioscience, Biotechnology, and Biochemistry; 56: 1541-1545: (1992).
CrossRef - Balti R, Bougatef A, Sila A, Guillochon D, Dhulster P, Nedjar-Arroume N. Nine novel angiotensin I-converting enzyme (ACE) inhibitory peptides from cuttlefish (Sepia officinalis) muscle protein hydrolysates and antihypertensive effect of the potent active peptide in spontaneously hypertensive rats. Food Chemistry; 170: 519: (2015).
CrossRef - Ren Y, Wan D-G, Lu X, Chen L, Zhang T, Guo J-L. Isolation and characterization of angiotensin I converting enzyme inhibitor peptides derived from porcine hemoglobin. Scientific Research and Essays; 6(30): 6262-6269: (2011).
- Qian Z-L, Jung W-K, Lee S-H, Byun H-G, Kim S-K. Antihypertensive effect of an angiotensin I-converting enzyme inhibitory peptide from bulfrog (Rana catesbeiana Shaw) muscle protein in spontaneously hypertensive rats. Process Biochemistry; 42: 1443-1448: (2007).
CrossRef - Yano S, Suzuki K, Funatsu G. Isolation from alpha-zein of thermolysin peptides with angiotensin I-converting enzyme inhibitory activity. Bioscience, Biotechnology, and Biochemistry; 60(4): 661-663: (1996).
CrossRef - Nakahara T, Sano A, Yamaguchi H, Sugimoto K, Chikata H, Kinoshita E, Uchida R. Antihypertensive effect of peptide-enriched soy sauce-like seasoning and identification of its Angiotensin I-Converting Enzyme inhibitory substances. Journal of Agricultural and Food Chemistry; 58: 821-827: (2010).
CrossRef - van Platerink C. J, Janssen H-G. M, Haverkamp J. Application of at-line two-dimensional liquid chromatography-mass spectrometry for identification of small hydrophilic angiotensin I-inhibiting peptides in milk hydrolysates. Analytical and Bioanalytical Chemistry; 391: 299-307: (2008).
CrossRef - Cheung H-S, Wang F-L, Ondetti M. A, Sabo E. F, Cushman D. W. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. The Journal of Biological Chemistry; 255: 401-407: (1980).
- Meisel H. Overview on milk protein-derived peptides. International Dairy Journal; 8:363-373 (1998).
CrossRef - Gomez-Ruiz J. A, Ramos M, Recio I. Identification of novel angiotensin-converting enzyme-inhibitory peptides from ovine milk proteins by CE-MS and chromatographic techniques. Electrophoresis; 28: 4202-4211: (2007).
CrossRef - Mine Y, Shahidi F. ACE inhibitory peptides in: Nutraceutical proteins and peptides in health and disease. CRC Taylor & Francis Group, Boca Raton, London, New York; 269-315: (2006).
- Sentandreu M. A., Toldra F. Evaluation of ACE inhibitory activity of dipeptides generated by the action of porcine muscle dipeptidyl peptidases. Food Chemistry; 101: 1629-1633: (2007).
CrossRef - Liu R, Zheng W, Li J, Wang L, Wu H, Wang X, Shi L. Rapid identification of bioactive peptides with antioxidant activity from the enzymatic hydrolysate of Mactra veneriformis by UHPLC–Q-TOF mass spectrometry. Food Chemistry; 167: 484-489: (2015).
CrossRef - Yokomizo A, Takenaka Y, Takenaka T. Antioxidative activity of peptides prepared from Okara protein. Food Science and Technology Research; 8(4): 357-359: (2002).
CrossRef - Chen H-M, Muramoto K, Yamauchi F. Structural analysis of antioxidative peptides from soybean b-conglycynin. Journal of Agricultural and Food Chemistry; 43: 574-578: (1995).
CrossRef - Sagardia I, Roa-Ureta R. H, Bald C. A new QSAR model, for angiotensin I-converting enzyme inhibitory oligopeptides. Food Chemistry; 136: 1370–1376: (2013).
CrossRef - Wu J, Aluko R. E, Nakai S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure-Activity Relationship Modeling of Peptides Containing 4-10 Amino Acid Residues. QSAR & Combinatorial Science; (10): 873 – 880: (2006).
CrossRef - Wu J, Aluko R. E, Nakai S. Structural requirements of angiotensin I converting enzyme inhibitory peptides: Quantitative structure and activity relationship study of di and tripeptides. Journal of Agricultural and Food Chemistry; 54(3): 732–738: (2006).
CrossRef - Foltz M, van Buren L, Klaffke W, Duchateau G.S. Modeling of the Relationship between Dipeptide Structure and Dipeptide Stability, Permeability, and ACE Inhibitory Activity. Journal of Food Science; 74(7): 243-51: (2009).
CrossRef - Guang C. E, Phillips R. D. Performance of four sets of amino acid descriptor scales for evaluating quantitative structure-activity relationships of angiotensin I converting enzyme inhibitory dipeptides. Journal of Food Agriculture and Environment; 10(2): 227-232: (2012).
- Cheung H. S, Wang F. L, Ondetti M. A, Sabo E. F, Cushman D. W. Binding of peptide-substrates and inhibitors of angiotensin-converting enzyme– Importance of the COOH-terminal dipeptide sequence. The Journal of Biological Chemistry; 255(2):401–407: (1980).
- Tian M, Fang B, Jiang L, Guo H, Cui J. Y, Ren F. Structure-activity relationship of a series of antioxidant tripeptides derived fromβ-Lactoglobulin using QSAR modeling. Dairy Science & Technology; 95: 451–463: (2015).
CrossRef - Li Y. W, Li B. Characterization of structure-antioxidant activity relationship of peptides in free radical systems using QSAR models: key sequence positions and their amino acid properties. Journal of Theoretical Biology; 318: 29–43: (2013).
CrossRef - Davies M. J. Protein oxidation and peroxidation. Biochemical Journal; 473: 805–825: (2016).
CrossRef - Nakamura Y, Yamamoto N, Sakai K, Yamazaki S, Takano T. Purification and characterization of angiotensin I-converting enzyme inhibitors from sour milk. Journal of Dairy Science; 78: 777-783: (1995).
CrossRef - Murray B. A, FitzGerald R. J. Angiotensin converting enzyme inhibitory peptides derived from food proteins: biochemistry, bioactivity and production. Current Pharmaceutical Design; 13: 773-791: (2007).
CrossRef - Kim S. K., Byung H.G., Park P.J., Shahidi F. Angiotensin I converting enzyme inhibitory peptides purified from bovine skin gelatin hydrolysate. Journal of Agricultural and Food Chemistry; 49: 2992-2997: (2001).
CrossRef - Wu H-C, Shiau C-Y, Chen H-M, Chiou T-K. Antioxidant activity of carnosine, anserine, some free amino acids and their combinations. Journal of Food and Drug Analysis; 11: 148-153: (2003).





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








