Inhibition of Enzymatic Browning by Onion (Allium Cepa L.): Investigation on Inhibitory Mechanism and Identification of Active Compounds
 , Sukarno1*, Nancy Dewi Yuliana1, Slamet Budijanto1
, Sukarno1*, Nancy Dewi Yuliana1, Slamet Budijanto1 1Department of Food Science and Technology, Bogor Agricultural University, Bogor, Indonesia
2Department of Fisheries Extension, Jakarta Fisheries University, Jakarta, Indonesia
Corresponding Author Email: dsukarno@apps.ipb.ac.id
DOI : http://dx.doi.org/10.12944/CRNFSJ.6.3.19
Download this article as:
![]()
Presence of browning or black-spot in fresh foods can adversely affect consumer acceptance. Onion has been reported to exert inhibitory activity against browning reaction. This research aimed to uncover the mechanism and identify active compounds in onion responsible for PPO inhibitors based on metabolomic approach. Onion was fractioned using different solvents, i.e n-hexane; chloroform; ethyl acetate; water, respectively. As a result, ethyl acetate fraction (EAF) of the onion demonstrated the strongest inhibition to PPO in comparison with other fractions, i.e. n-hexane, chloroform, and water. The reversible inhibitory activity of PPO by EAF occurred with presence of L-DOPA as substrate through competitive inhibition and Cu chelation in the active side of the PPO. Based on 1H-NMR (X) score plot and PPO inhibition (Y) using OPLS, NMR signals revealed that active compounds accounting for inhibition of PPO included quercetin, kaempferol, cyanidin 3.4’-di-O-β-glucopyranoside, quercetin 4-O-β-D-glucopyranoside, cyanidin 7-O-(3”-O-glucosyl-6”-O-malonyl-β-glucopyrano-side)-4’-O-β-glucopyranoside, cyanidin 3-(6”-O-malonyl) laminaribioside’.
KEYWORDS:Cyaniding; Kaempferol; Metabolomic; Onion; PPO Inhibitor; Quercetin
Introduction
Browning or black-spot constitutes a post-harvest problem occurring in fresh foods. It often exists in fresh-cut fruits and vegetables. However, black-spot is also present in crustacean such as shrimp during post-mortem process, as a result of enzymatic reaction by PPO responsible for formation of black pigment. Although the pigment is not deleterious to health, its presence on fresh shrimp closely relates to low consumer acceptance.1
PPO enabled to catalyze reaction of monophenol hydroxylation to form o-diphenol through monophenol oxidase action and oxidize o-diphenol into o-quinon through diphenolase activities. Quinone is a reactive compound and spontaneously able to form polymers with quinone itself or with other aromatic compounds, yielding a black melanin polymer. Monophenol oxidase is also recognized as laccase (EC 1.10.3.2), while diphenol oxidase is known as catechol oxidase (EC 1.10.3.1). PPO is commonly present as tyrosinase (EC 1.14.18.1) that can use two substrates, i.e. monophenol and diphenol such as mushroom tyrosinase. The PPO enzyme nomenclature seems to be inconsistent, depending on types of substrate, i.e. tyrosinase, phenolase, catechol oxidase, catecholase, o-diphenol oxidase, monophenol oxidase, and cresolase. Structurally, there are six histidine residues liganded to the copper of active site of PPO. The enzyme may occur in many sources including bacteria, animals, and plants.2
Furthermore, flavonoid is also reported to exert anti-browning activity. Additionally, depigmentation of melanin by polyphenolic was also regarded as anti-browning activity.3 Attempts to deal with browning or black-spot closely relate to antioxidant activity4 since melanin is a product of enzymatic oxidation. However, natural bioactive compounds demonstrating antioxidative properties are not always associated with browning inhibition.
Onion is rich in phenolic and flavonoid, and has a high antioxidant activity. Previous study reported that some beneficial compounds including gallic acid, ferulic acid, quercetin, and proanthocyanin, were detected in onion studied.5 Furthermore, a study showed inhibitory effect of onion extract on the enzymatic browning of an edible yam (Dioscorea cayenensis-rotundata cv. Kponan).6 Nevertheless, the mode of action and compounds responsible for the browning inhibition has remained unclear.
Scientific evidence related to mechanism and the role of active compounds is essential to investigate. For this reason, isolation and metabolomic approach are feasible method to determine the active compounds. Metabolomics assay enables to identify active compounds quantitatively and qualitatively present in a material. Even though using a small amount of sample, the approach is capable of producing comprehensive comparison according to the activity of the compounds, without the need for isolating the active principles.7 This study aimed to uncover the plausible mechanism and identify active compounds in onion as PPO inhibitor using metabolomics approach.
Materials and Methods
Onion was obtained from Brebes, Central Java. Chemicals were analytical grade, i.e. methanol, n-hexane, chloroform, ethyl acetate, NaH2PO4, K2HPO4, NaNO2, NaOH, L-DOPA (Merck, USA), quercetin (Merck, HPLC Grade, USA). Commercial PPO enzyme (EC.1.14.18.1, 5571 U/mg 4.3 mg) was purchased from Sigma Chemical Co (St. Louis. MO), while sodium metabisulphite (SMS) as positive control of PPO inhibitor, was obtained from local chemical supplier.
Fractionation of Onion8
Fresh onion (800g) was washed, freeze-dried for 24 h, and powdered. The dried onion powder (100 g) was extracted with 80% methanol (500 mL). The mixture was sonicated using ultrasonic bath (Bransonic Ultrasonic Cleaner model 8510E MTH, USA) for 30 min, and the supernatant was collected. The the supernatant was dried using rotary evaporator (Butchi Rotavapor R-210, BÜCHII, Labortechnik, Switzerland) at 45 °C. About 10 g methanolic onion extract was dissolved in distilled water (250 mL) and n-hexane (250 mL) filled to separatory funnel. The funnel was shaken vigorously and it was set at room temperature until two distinct layers were observed. The n-hexane fraction was separated. The fractionation was also performed using similar aforementioned protocol using other solvents, i.e. chloroform and ethyl acetate, in five replications for each fraction. All fractions were evaporated to obtain the dry fraction.
PPO Inhibition Test9
PPO enzyme solution (400 µL, 23 Unit/mL) was added with 400 µL of onion fraction (dry basis, 10 mg/mL) and incubated for 30 min. The mixture (200 µL) was taken and added with 15 mM L-DOPA (600 µL), 50 mM buffer phosphate (400 µL, pH 6.0), distilled water (1000 µL), and then incubated for 3 min at 45 °C. The formation of dopachrome was spectrophotometrically checked at wavelength of 475 nm using UV-2425 spectrophotometer (Shimadzu, Kyoto, Japan). One unit of PPO was defined as the increment of absorbance by 0.001/min/mL at A475. The inhibitory activity was expressed as the percentage inhibition as follows:
% of PPO inhibition = {(A-B)/A} x 100%
Where A: PPO activity for control (absence of fraction), B: PPO activity with presence of fraction.
Reaction Kinetics of Onion Fractions 10
Lineweaver-Burk plot was made by plotting rate of reaction and substrate concentration [S] with the equation as follows: 1/ = .maks × 1/[] + 1/maks. Linear regression model was used to determine constant Michaelis-Menten and maximum velocity max. The plot could differentiate performance of selected onion fraction as competitive inhibitor, non-competitive inhibitor, or uncompetitive. The solution of selected onion fraction (400 L) was made at different concentrations: 0.25; 0.5; 1 mg/mL in tube and added with 400 L of PPO enzyme solution (0; 0.25; 0.5; 1 mg/mL). The L-DOPA as a substrate was made at concentration of 0.1, 0.25, 0.5, 1, 1.5, 2 mg/mL in buffer solution of sodium phosphate at pH 6.0.
Electrophoresis Experiment11
Native polyacrylamide gel electrophoresis (PAGE) experiment was applied to investigate the effect of onion fraction on PPO activity. PPO, onion fraction and phosphate buffer solution (50 mM, pH 6.0) were mixed in a vial tube. The fraction was dissolved in methanol 0.5% and dissolved in water at concentration of 1, 5, 10 mg/mL, sodium metabisulphite 1.25%. Afterwards, the solution (100 µL) was mixed with PPO (100 µl), and kept for 30 min. A mixture comprised (20 µL) of bromofenol blue (0.001%, w/ v) and glycerol (0.05%, v/v) was injected to each well to control the process. Electrophoresis experiment was performed at 4 °C using Mini PROTEAN system (Bio-Rad, Hercules, CA), with Tris–Glicine (pH 8.3) as running buffer. The gel was then soaked in a solution containing L-DOPA (15mM).
Wavelength Scanning 10
The chelation of copper from the active site of PPO was evaluated using wavelength scanning by spectrophotometer (UV Vis-2425, Shimadzu). Distilled water (2.8 mL) was added with selected onion fraction (0.1 mL) previously dissolved in methanol 0.5%. The mixture was transferred to 3 tubes containing PPO (tube 1), 0.1 mL of CuSO4 (tube 2), and 0.1 mL of buffer sodium phosphate pH 6.0 (tube 3). Ultimately, the solution was spectrophotometrically scanned at wavelength of 200-600 nm, while distilled water was used as reference.
Compound Identification by 1H-NMR and 2D NMR J-Resolved
Active compounds responsible for PPO inhibition were identified using 1H-NMR and 2D NMR at 500 MHz using JEOL type ECZR Spectrometer = JNM-ECZ500R/S1 (Japan), with functional frequency of 500.15991521 MHz at room temperature (20.1 °C). A 10 mg fraction of onion was dissolved in 1 mL CH3OH-d4 (MeOD). The mixture was homogenized in a vortex for 1 min, ultra-sonicated for 15 min, then centrifuged at 13,000 rpm for 10 min in NMR tube was contained this supernatant (0.6 mL) and analyzed to 1H-NMR. All protons in one dimension or single pulse identified as axis X. 1H-NMR spectrum of each sample was required 1.74587904 s, that consists of 128 scans and a width of 5 ppm. The fraction which had the most active signal were selected and continued to be analyzed in 2D NMR using Homo J-resolved. Spectrum of J-resolved was needed 50 min which X accuracy duration 0.65404928 s, Y accuracy time = 2.56 s containing 16 scans per 2048 increase for the spin-spin axis coupling constant. The spectral widths were 50 Hz and 8 K for the chemical shift axis with spectral widths of 500 Hz. Time for relaxation delay was 1.5 in 20.7 ˚C.
Multivariate Data Analysis7
JDF files binned spectra in the same parameters. All spectra included a spectra width (δ 0.04) set up an area of 0.5-10 ppm to reach a total 230 integrated areas per NMR spectrum using MestReNova version 12-0.2.20910 (Mestrelab Research, S.L , Spain). OPLS analysis was conducted using SIMCA –P Software (v.12.0, Umetrich, Umea, Sweden) based on the pareto method of scaling. The quality of the model was described with the model accuracy criteria (R2Y) and prediction accuracy (Q2Y) and validated by CV ANOVA and permutation test.
Results and Discussion
Fractionation of Onion with Various Solvents
The results showed that ethyl asetate fraction (EAF) of the onion demonstrated the highest inhibition against PPO (90.33 ± 0.93) in comparison with the onion fractioned by other solvents, i.e. n-hexane (60.13 ± 0.78), chloroform (52.97 ± 0.34), and water (47.09 ± 0.90) (Figure 1). This suggests that solvent for fractionation is considered as a critical factor affecting PPO inhibitory activity. The solvent may alter the fractionability of phytochemical compounds, thus resulting in different effects on PPO inhibition. The difference in PPO inhibitory activity may also result from extraction temperature and solvent.13 Different polarity of the solvents (binary system), for instance methanol and water at a certain ratio (70%), enabled to extract more phenolic and flavonoid compounds in comparison with the use of single solvents.14
We found that EAF exhibited higher inhibitory activity than water extract of ‘Recas’ onion, and could attenuate PPO activity in avocado fruits at 89.71% 15. EAF was also found to exert stronger effect on PPO inhibition than heated onion juice, and demonstrate anti-browning activity through inhibiting PPO activity in ‘Birgah’ eggplant at 54.2%.16
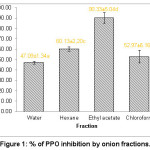 |
Figure 1: % of PPO inhibition by onion fractions. |
Reaction Kinetics of EAF
As a result, the increase in EAF concentration could reduce slope of reaction velocity (V) vs PPO concentration. This indicates that EAF showed a reversible inhibition towards PPO activity in presence of substrate L-DOPA (Figure 2). Natural extract often occurs as reversible inhibitor. The PPO possesses a stronger affinity to substrate L-isomers such as L-DOPA than D-isomer, and also has various specific substrates.17
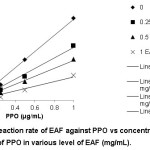 |
Figure 2: Reaction rate of EAF against PPO vs concentration of PPO in various level of EAF (mg/mL). |
In our experiment, Lineweaver-Burk plot was established to examine the inhibition mechanism. The concentration of EAF (0, 0.25, 0.5, 1 mg/mL) positively correlated with Km value in presence of substrate L-DOPA, resulting in Km value of 0.244, 0.956, 1.481, 2.106 mM, respectively. Km constitutes a Michaelis-Menten constant, which represents the remaining substrate after the enzyme reached a half of maximum rate Vmax. In this case, Vmax was found at 0.0164 mM-1 min, and it was unaltered with the increasing in EAF concentration. It means that the maximum substrate concentration (V max) was replaced the position by EAF in the active side of the PPO enzyme. As depicted in Figure 3, we demonstrated that EAF showed a competitive inhibition. The competitive inhibitor is a substance that binds with free enzyme, thus preventing substrate from binding active site in enzyme.17 The hydroxyl groups of inhibitor could bind active site of PPO, changing the enzyme conformation.18
For competitive inhibition, the inhibition constant KI was assessed using Kmapp = Km [1 + ([I]/KI)], where Kmapp is the apparent Km for the reaction due to presence of EAF. The constant KI was determined by plotting Michaelis-Menten constant (Kmapp) and EAF as inhibitor, resulting in a linear line.
 |
Figure 3: Lineweaver–Burk plot describing reaction between L-DOPA and PPO with various concentrations of EAF. |
As a result, constant Km was 0.4038 mM, while Km/KI was 1.8129. Based on this calculation, the constant KI was 0.4038 mM/1.8129 or 0.222 mM. The action of inhibitor accounted for a higher Km’ value than initial Km. The constant KI demonstrates the enzyme affinity for a substrate. In this case, a greater Km means a lower enzyme affinity. The presence of inhibitor induced enzyme to reach maximum reaction rate, but required either longer time or more substrate.19
In the case of anti-browning activity, previous studies have reported the competitive inhibition induced by compounds such as salidroside which is a flavonoid from Rhodiola rosea10 and catechin from greentea.20 Meanwhile, anti-browning activity on edible yam (Dioscorea cayenensis-rotundata cv. Kponan) by water extract of onion and garlic was found to exert various types of inhibition, i.e. non-competitive (yellow and green onion extract), competitive (red, small, and white onion extract), and uncompetitive (garlic extract).6
Electrophoresis Experiment of EAF
Figure 4 shows the result of native polyacrylamide gel electrophoresis (PAGE) experiment demonstrating the inhibitory effect of EAF on PPO activity. Native PAGE is performed in the absence of sodium dodecyl sulphate (SDS) which is frequently applied to disrupt the structure of protein and promote reduction on buffer system. It is also applicable for determination of molecular weight since the electrophoretic mobility of protein molecules relies on their molecular mass. In the case of native PAGE, the molecular mobility is dependent on protein charge and hydrodynamic size. The electrical charge during electrophoresis is mainly affected by composition of amino acids and pH of the running buffer system. Native PAGE is meaningful for discovering functional properties of the enzyme, including effect of enzyme inhibitor.21
In our experiment, Rf value of PPO was 0.44. The similar Rf value was applied in another study using mushroom tyrosinase, investigating the plausible mechanism of enzymatic browning inhibition by NaCl.11 Rf is migration distance of the enzyme divided by length of electrophoretic gel. Using similar gel composition and running time, the same PPO possesses a similar value of Rf.22
A remarkable change in the enzyme occurred in presence of EAF, accounting for formation of dark color. This suggests that EAF provides a noticeable impact on reduction of melanin. This inhibitor could bind PPO, thus preventing the enzyme from interacting with substrate L-DOPA. The increase in EAF concentration (0, 1, 5, 10 mg/mL) resulted in a thinner dark band. As exhibited in Figure 4, the strongest inhibitory effect was found at EAF concentration of 10 mg/mL, yielding the thinnest band, which is similar to that treated with SMS of 12.5 mg/mL.
 |
Figure 4: The changes in PPO after 30 min incubation with variouslevels of EAF observed using Native PAGE (lane 1-4 indicates10, 5, 1, 0 mg/mL of EAF, respectively, lane 5 is 12.5 mg/mLf SMS. L-DOPA is used as substrate). |
Wavelength Scanning
The absorption peak shifts from 390 nm to 440 nm, which indicates Cu can react with EAF (Figure 5). EAF could be also meaningful as metal chelator, thus making it noticeable as PPO inhibitor due to presence of Cu in active site of the enzyme. Flavonoid compound was also reported to display antioxidant activities against redox metal (Cu, Fe, and Zn) ions.23
Previously, kojic acid was also reported to have inhibitory effect on PPO and metal chelator.24 Some PPO inhibitor demonstrating competitive mechanism could serve as metal chelator, non-metabolite analog, and/or derivate from the real substrate.25 The molecular structure of cyanidin belonged to polyphenolic compounds is responsible for metal-chelating properties, i.e. ortho-hydroxy groups in the B-ring, 2,3-double bond in the conjugation, and 4-oxo-function in the C-ring. Flavonoid enables to form complex with Cu2+ ions in 3- or 5- hydroxyl and 4-ketosubstituent or hydroxyl groups in ortho position of the B-ring.26
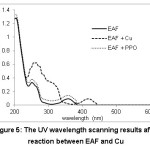 |
Figure 5: The UV wavelength scanning results after reaction between EAF and Cu |
Identification of Compounds Using Metabolomics Approach
The active compounds responsible for PPO inhibition were identified using metabolomics approach. The method is essential to simplify isolation stages, but requires the use of appropriate multivariate analysis due to a large quantity of data. The 1H-NMR analysis results in chemical shift for each fraction (4 fraction) of onions (5 replications that data is made on average as shown in Figure 1) used to estimate the active compounds.
Metabolomics data sets are large, thereby requiring multivariate analysis such as Orthogonal Projections to Latent Structures (OPLS). OPLS is applied to interpret data through combining them after screened using Orthogonal Signal Correction (OSC) to produce PLS model, to effectively separate Y-predictive variation from Y-uncorrelated variation in X-axis.27
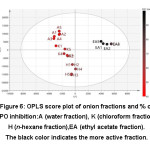 |
Figure 6: OPLS score plot of onion fractions and % of PPO inhibition:A (water fraction), K (chloroform fraction), H (n-hexane fraction),EA (ethyl acetate fraction). The black color indicates the more active fraction. |
We performed metabolomics analyses to explore the relationship between the metabolome and bioactivity of diverse onion fractions. A good separation between lower activity fractions and higher activity fraction was showed by the OPLS score plot (Figure 6). The OPLS value of R2X, R2Y, and Q2 of this model is 0.903, 0.936, and 0.774, respectively, indicating the reliability of the model. Thus, OPLS may be used further to identify chemical shifts which can be attributed to the active compounds.
Metabolomics data model is unacceptable when Q2 value was less than 0 or Q2(cum) of < 0.05 and > 2. Q2 > 40% and R2 > 50% was generally accepted as being indicative of a robust model, although these values need to be confirmed for the bioactivity condition.28
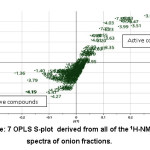 |
Figure 7: OPLS S-plot derived from all of the 1H-NMR spectra of onion fractions. |
OPLS S-plot was used to to determine the NMR chemical shift (ppm) which have a positive correlation with the activity (Figure 7). By using X variant plot (not shown), a fraction having abundant NMR signal of the respective chemical shifts was then measured for 2D NMR J-resolved (supplementary information), through comparison with previously reported NMR data of compounds identified from onion and other plants.29 After determination of the chemical shift with the most inhibitory activity, further comprehensive investigation was conducted to identify the compound using 2D NMR in order to untangle NMR overlapping signals (Figure 8). The 2D NMR analysis was performed to determine spin-spin coupling that may display magnetization transfer among molecules.30 J-resolved spectroscopy constitutes one of the 2D NMR tests that could be carried out easily and accurately in determining 2D pulse sequence. Double-Spin Echo variant (DSE JRES) is applied to reduce artifact as a consequence of strong coupling. Water suppression was reduced using WET, and followed by presaturation. WET is employed to remove water interruption that adversely affects the sensitivity of instrument and 2D NMR analysis results. The single-channel detection was applied, similar to that for homonuclear J-resolved. At first, coupling was reduced, while strongly coupled proton could be used as data.31 The 2D NMR is to define coupling constant (J) of hydrogen atom derived from 1H-NMR signal. Additionally, metabolites at lower levels and with difficulties on the signal detection required 1H-NMR spectra for identification.32
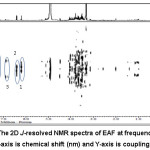 |
Figure 8: The 2D J-resolved NMR spectra of EAF at frequency of 500 MHz. The X-axis is chemical shift (nm) and Y-axis is coupling constant. |
The results of 1H and 2D NMR analysis are chemical shift, splitting pattern, and J-coupling constants of the EAF metabolite that shows inhibitory activity towards PPO (Table 1). The collected data were then compiled with those from previous studies on similar plant, thus information related to compounds contributing to inhibitory activity towards PPO was obtained. In this experiment, other compounds present in EAF were also detected at chemical shift of +4.2-3.5 ppm, including glycoprotein and amino acids, but they did not correlate with the anti-browning activity. Furthermore, aliphatic chemical shift was found at 4.5-0.7 ppm, while methylene and aromatic group or phenolic compounds were observed at chemical shift of 1.6 ppm and 6-8 ppm, respectively.33
Table 1: The 1H 2D NMR Spectra of EAF
| No. | Chemical shift (ppm) | Splitting Pattern (J) | Metabolite |
| 1 | 6.166.37 | d (J =2.5)d (J=2) | Quercetin34 |
| 2 | 6.85 | d (J=8) | Kaempferol34 |
| 3 | 7.61 | d (J=10) | Cyanidin 3.4’-di-O-β-glucopyranoside35 |
| 4 | 7.70 | d (J=2) | Quercetin 4 -O- β-D-glucopyranoside34 |
| 5 | 7.73 | d (J=2) | Cyanidin 7-O-(3”-O-glucocyl-6”-O-malonyl-b-glucopyranoside)-4’-O-b-glucopyranoside34 |
| 6 | 7.87 | d (J=2) | Cyanidin 3-(6″-O-malonyl) laminaribioside’36 |
The solvent for fractination of onion seemed to remarkably affect the types of fractinationed flavonoid. The water extract of onion was reported to contain alkaloid, carbohydrate, reducing sugar, flavonoid, and glycoside, while tannin and phenolic compounds were found in the ethanolic extract. In both extracts, some components were also detected, including saponin, amino acid, protein, terpenoid, and steroid, but cardiac glucoside was not present.37 In the case of EAF, flavonoid compounds were detected at high abundance, primarily from flavonols such as kaempferol, quercetin, and quercetin glycoside (isoquercitrin, quercitrin and rutin).38
We found that phytochemical components in either fraction are diverse. Some polyphenol compounds in plant extract may provide synergistic effect.39 However, in a particular case, bioactivity of a single compound may be more powerful in comparison with other plant extracts, such as mimosine present in Leucaena leucocephala.9 Quercetin and kaempferol have been extensively researched for their anti-browning and antioxidant activity.40 However, investigation on anti-browning effect of cyanidin is rather scarce.
Presence of quercetin, kaempferol and cyanidin in onion fractions noticeably contributed to the inhibitory activity towards PPO. The PPO inhibitor could be either a compound or combination of compounds that naturally existed and accounted for formation of reversible binding and reduction of enzyme catalytic activity. Furthermore, phenolic compounds are also found to be PPO inhibitor, depending on availability and position of subsistent. The inhibitory properties towards PPO from polyphenolic compounds, e.g. quercetin and kaempferol (as flavonoid group), are associated with the existence of phenol and pyrene groups.41
Conclusion
Inhibitory mechanism of active compounds in onion towards PPO has been successfully investigated. Onion was fractioned using various solvents with different polarity (ethyl acetate, n-hexane, chloroform, and water), resulting in different inhibitory activities. Ethyl acetate fraction (EAF) showed the strongest inhibition against PPO with a reversible mechanism, in which L-DOPA was used as substrate. Enzyme kinetics demonstrated that type of PPO inhibition by EAF was competitive. The fraction was also able to form chelate complex with Cu in active site of PPO.
Metabolomics approach using 1H-NMR and 2D NMR instrument was employed in order to identify compounds that associated with PPO inhibition. As a result, flavonoid group was concluded to be contributor for the inhibitory activity. Quercetin and kaempferol have been widely recognized for their inhibitory activity towards PPO, but studies on the anti-browning activity by cyanidin have remained scarce. Therefore, further research is needed focusing particularly on the role of cyanidin in inhibition of browning and its potential antagonistic or synergistic interaction with quercetin and kaempferol.
Acknowledgements
The authors expressed sincere gratitude to the Marine and Fisheries Education Center (PUSDIK KP), the Ministry of Marine Affairs and Fisheries, Republic of Indonesia, for the financial support.
References
- Senapati S. R., Kumar G. P., Singh C. B. et al., Melanosis and quality attributes of chill stored farm raised whiteleg shrimp ( Litopenaeus vannamei ). J Appl Nat Sci. 2017;9(1):626-631.
CrossRef - Mishra B. B., Gautam S. Polyphonel Oxidases: Biochemical and Molecular Characterization, Distribution, Role and its Control. Enzym Eng. 2016;05(01):1-9. doi:10.4172/2329-6674.1000141
CrossRef - Fu B., Li H., Wang X., Lee F. S. C., Shufen Cui. Isolation and Identification of Flavonoids in Licorice and a Study of Their Inhibitory Effects on Tyrosinase. J Agric Food Chem. 2005;53(19):7408–7414. doi:doi: 10.1021/jf051258h
CrossRef - Wu L., Jou A., Chen S. et al., Antioxidant, Anti-inflammatory and Anti-browning Activities of Hot Water Extracts of Oriental Herbal Teas. Food Funct. 2010;1(2):200-208. doi:DOI: 10.1039/c0fo00047g
CrossRef - Cheng A., Chen X., Jin Q., Wang W., Shi J. Comparison of Phenolic Content and Antioxidant Capacity of Red and Yellow Onions. Czech J Food Sci. 2013;31(5):501-508.
CrossRef - Yapi J. C., Gnangui S.N., Dabonné S. Inhibitory Effect of Onions and Garlic Extract on the Enzymatic Browning of an Edible Yam ( Dioscorea cayenensis-rotundata cv . Kponan ) cultivated in Côte d Ivoire. Int J Curr Res Acad. 2015;3(1):219-231.https://doi.org/10.1016/j.jep.2013.08.012
- Kim H. K., Choi Y. H., Verpoorte R. NMR-based metabolomic analysis of plants. Nat Protoc. 2010;5(3):536-549. doi:10.1038/nprot. 2009;237.
- Sri Widyawati P., Budianta T. D. W., Kusuma F. A., Wijaya E. L. Difference of Solvent Polarity to Phytochemical Content and Antioxidant Activity of Pluchea indicia Less Leaves Extracts. Int J Pharmacogn Phytochem Res. 2014;6(4):850-855.
- Nirmal N. P., Benjakul S. Inhibition of melanosis formation in Pacific white shrimp by the extract of lead ( Leucaena leucocephala ) seed. Food Chem. 2011;128(2):427-432. doi:10.1016/j.foodchem.2011.03.048
CrossRef - Zhu Y., Chen C., Zhao S. et al., Inhibitory Mechanism of Salidroside on Tyrosinase. J Food Nutr Res. 2014;2(10):698-703. doi:10.12691/jfnr-2-10-8
CrossRef - He Q., Luo Y., Chen P. Elucidation of The Mechanism of Enzymatic Browning Inhibition by Sodium Chlorite. Food Chem. 2008;110:847-851. doi:10.1016/j.foodchem.2008.02.070
CrossRef - Yuliana N. D., Budijanto S., Verpoorte R., Choi Y. H. NMR metabolomics for identification of adenosine A1 receptor binding compounds from Boesenbergia rotunda rhizomes extract. J Ethnopharmacol. 2013. doi:10.1016/j.jep.2013.08.012
CrossRef - Sharif A., Saim N., Jasmani H., Ahmad W. Effects of solvent dan temperatur on extraction of colorant from onion (Allium cepa) skin using pressurized liquid extraction. Asian J Aplied Sci. 2010;3(4):261-268.
- Singh V., Krishan P., Shri R. Extraction of antioxidant phytoconstituents from onion waste. J Pharmacogn Phytochem. 2017;6(1):502-505.
- Roldan E., de Ancos B., Sancez-Moreno C., Cano M. P. Characterisation of Onion (Allium cepa L .) by-products as Food Ingredients with Antioxidant and Antibrowning Properties. Food Chem. 2008;108:907-916. doi:10.1016/j.foodchem.2007.11.058
CrossRef - Barbagallo R. N., Riggi E., Avola G., Patanè C. Biopreservation of “Birgah” Eggplant from Polyphenol Oxidase Activity Assayed In Vitro with Onion ( Allium Cepa L .) by-products. Chem Eng Trans. 2012;27(1984):1-6.
- Loizzo M. R., Tundis R., Menichini F. Natural and Synthetic Tyrosinase Inhibitors as Antibrowning Agents: An Update. Compr Rev Food Sci Food Saf. 2012;11(4):378-398. doi:10.1111/j.1541-4337.2012.00191.x
CrossRef - Taherkhani N., Gheibi N. Inhibitory Effects of Quercetin and Kaempferol as two Propolis Derived Flavonoids on Tyrosinase Enzyme. Biotechnol Heal Sci. 2014;1(2):1-5. doi:10.17795/bhs-22242
CrossRef - Gacche R. N., Shete A. M., Dhole N. A., Ghole V. S. Reversible inhibition of polyphenol oxidase from apple using L-cysteine. Indian J Chem Technol. 2006;13(5):459-463.
- Sae-Leaw T., Benjakul S., Simpson B. K. Effect of Catechin and Its Derivatives on Inhibition of Polyphenoloxidase and Melanosis of Pacific White Shrimp. J Food Sci Technol. 2017;54(5):1098-1107. doi:10.1007/s13197-017-2556-1
CrossRef - Nowakowski A. B., Wobig W. J. Petering D. H. Native SDS-PAGE: High Resolution Electrophoretic Separation of Proteins With Retention of Native Properties Including Bound Metal Ions. Metallomics. 2015;6(5):1068-1078. doi:10.1039/c4mt00033a.Native
- Mathews H. M., Moss D. M., Healy G. R., Visvesvara G. S. Polyacrylamide gel electrophoresis of isoenzymes from Entamoeba species. J Clin Microbiol. 1983;17(6):1009-1012.
- Cherrak S. A., Mokhtari-Soulimane N., Berroukeche F. et al., In Vitro Antioxidant versus Metal Iion Chelating Properties of Flavonoids: A Structure-Activity Investigation. PLoS One. 2016;11(10):1-21. doi:10.1371/journal.pone.0165575
CrossRef - Hashemi S. M., Emami S. Kojic acid-derived tyrosinase inhibitors: synthesis and bioactivity. Pharm Biomed Res. 2015;1(1):1-17. doi:10.18869/acadpub.pbr.1.1.1
CrossRef - Chen K., Zhao D. Y., Chen Y. L. et al., A Novel Inhibitor Against Mushroom Tyrosinase with a Double Action Mode and Its Application in Controlling the Browning of Potato. Food Bioprocess Technol. 2017;10(12):2146-2155. doi:10.1007/s11947-017-1976-2
CrossRef - Nimse S. B., Pal D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015;5(35):27986-28006. doi:10.1039/c4ra13315c
CrossRef - Worley B., Powers R. Multivariate Analysis in Metabolomics Bradley. Curr Metabolomics. 2015;1(1):92-107. doi:10.2174/2213235X11301010092.Multivariate
- Blasco H., Błaszczyński J., Billaut J. C. et al., Comparative analysis of targeted metabolomics: Dominance-based rough set approach versus orthogonal partial least square-discriminant analysis. J Biomed Inform. 2015;53:291-299. doi:10.1016/j.jbi.2014.12.001
CrossRef - Li Z. Y., Ding L. L., Li J. M. et al., 1H-NMR and MS Based Metabolomics Study of the Intervention Effect of Curcumin on Hyperlipidemia Mice Induced by High-fat Diet. PLoS One. 2015;10(3):1-16. doi:10.1371/journal.pone.0120950
CrossRef - Bingol K., High N., Field M. Multidimensional Approaches to NMR-Based Metabolomics. Anal Chem. 2015;86(1):47-57. doi:10.1021/ac403520j.Multidimensional
- Guennec A Le. Fast 2D NMR Spectroscopy for Complex Mixtures. English: Ecole Polytechnique; 2015. https://pastel.archives-ouvertes.fr/tel-01191697%0D.
- Dona A. C., Kyriakides M., Scott F. et al., A Guide to The Identification of Metabolites in NMR-Based Metabonomics/Metabolomics Experiments. Comput Struct Biotechnol J. 2016;14:135-153. doi:10.1016/j.csbj.2016.02.005
CrossRef - Budantsev A. Y., Uversky V. N., Kutyshenko V. P. Analysis of the Metabolites in Apical Area of Allium cepa Roots by High Resolution NMR Spectroscopy Method. Protein Pept Lett. 2010;17(1):86-91.
CrossRef - Mohamed G. A. Alliuocide A: A New Antioxidant Flavonoid from Allium cepa L. Phytopharmacology. 2013;4(2):220-227.
- Fossen T., Slimestad R., Andersen Ø. M. Anthocyanins with 4 0 -Glucosidation from Red Onion , Allium cepa. Phytochemistry. 2003;64:1367-1374. doi:10.1016/j.phytochem.2003.08.019
CrossRef - Terahara N., Yamaguchi M., Honda T. Malonylated Anthocyanins from Bulbs of Red Onion, Allium cepa L. Biosci Biotechnol Biochem. 1994;58(7):1324-1325. doi:10.1271/bbb.58.1324
CrossRef - Vandana S., Ramesh K. In Vitro Antioxidant and Phytochemical analysis of Methanolic and Aqueous Extract of Allium cepa. Int J Recent Sci Res. 2017;8(11):21923-21928. doi:10.24327/IJRSR
CrossRef - Olayeriju O. S., Olaleye M. T., Crown O. O. et al., Ethylacetate Extract of Red Onion (Allium cepa L.) Tunic Affects Hemodynamic Parameters in Rats. Food Sci Hum Wellness. 2015;4(3):115-122. doi:10.1016/j.fshw.2015.07.002
CrossRef - De Freitas Formenton Macedo Dos Santos V. A., Dos Santos D. P., Castro-Gamboa I., Zanoni M. V. B., Furlan M. Evaluation of Antioxidant Capacity and Synergistic Associations of Quinonemethide Triterpenes and Phenolic Substances from Maytenus ilicifolia (Celastraceae). Molecules. 2010;15(10):6956-6973. doi:10.3390/molecules15106956
CrossRef - Farag M. A., Ali S. E., Hodaya R. H. et al., Phytochemical Profiles and Antimicrobial Activities of Allium cepa red cv. and A. sativum Subjected to Different Drying Methods: A Comparative MS-Based Metabolomics. Molecules. 2017;22(761):1-18. doi:10.3390/molecules22050761
CrossRef - Chang T. An Updated Review of Tyrosinase Inhibitors. Int J Mol Sci. 2009;10:2440-2475. doi:10.3390/ijms10062440
CrossRef
Scopus Citation(s): 11
Accepted on: 06-12-2018
Second Review by: Dr.Haripriya Shanmugam (India)
Final Approval by: Prof. Min-Hsiung Pan





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








