Antioxidant and Anti-inflammatory Properties of Prunella vulgaris Tea
Rachael Tan1
, and Wai Mun Loke2* 1School of Applied Sciences, Nanyang Polytechnic, Ang Mo Kio Ave 8 Singapore.
2Innovprof, 27 Orange Grove Road Singapore .
Corresponding Author Email: wai.mun.loke@innovprof.com
DOI : http://dx.doi.org/10.12944/CRNFSJ.10.2.9
Download this article as:
![]()
Prunella vulgaris is a traditional herb commonly consumed as brewed tea in Asian and European countries. The therapeutic antioxidant and anti-inflammatory activity of this traditional herbal tea are relatively unknown. The study examined, using established in vivo markers of oxidative stress and inflammation, the antioxidant and anti-inflammatory actions of Prunella vulgaris tea in-vitro, which were compared with those measured from green and black tea. Prunella vulgaris tea significantly decreased the human neutrophilic formations of F2-isoprostanes, lipid hydroperoxides, and leukotriene B4, while increasing those of nitrite and nitrate. The measured antioxidant and anti-inflammatory effects were greater than those from green and black tea. Prunella vulgaris tea exerted antioxidant effects via electron-transfer radical scavenging and up-regulated antioxidant enzyme activities, and anti-inflammatory effects by modulating 5-lipoxygenase, myeloperoxidase, and inducible nitric oxide pathways. The study results provide evidence supporting further investigations in the in vivo conditions.
KEYWORDS:Antioxidant; Anti-Inflammatory; Herbal Tea; Prunella vulgaris
Introduction
Oxidative stress and inflammation have been associated with the pathogenesis of cardiovascular diseases, such as atherosclerosis.1 Oxidative stress arises when the balance between the in vivo oxidants and antioxidants is distorted in favor of the oxidants.1 The excess oxidants damage essential proteins, fats, and nucleic acids, inflicting irreversible in vivo injuries.1 The overwhelmed antioxidants cannot counter these oxidative damages. Exogenous antioxidants may help prevent oxidative tissue damage by eliminating the excess oxidants and restoring the oxidative balance.2 Inflammation is an essential physiological response of the body to various pathological processes such as pathogen invasion and tissue injury.1 It involves infiltration and subsequent activation of the immune cells, such as blood neutrophils and monocytes, to the site of injury or pathogen infiltration and the production of inflammatory mediators such as chemokines and cytokines.1 Unresolved or overly expressed inflammation results in extensive tissue damage beyond the required immunity responses. Therapeutic intervention that alleviates oxidative stress and inflammation may delay the pathogenesis of cardiovascular and metabolic diseases.
Prunella vulgaris (PV) is a herb under the mint family commonly used in Asian and European countries. The name “Prunella” originates from the German word “Brunellen,” which translates to inflammation of the mouth, and “vulgaris” comes from a Latin adjective meaning “vulgar” or “common”. In Asian and European cultures, PV is traditionally consumed as brewed tea to relieve fevers, diarrhea, and sore mouth.3 In Southeast Asian cultures, the same herb has been traditionally consumed as “cooling tea,” which Traditional Chinese Medicine practitioners believed to remove the accumulated “heat”, restore the Ying-Yang balance of the human body, and maintain good health.3 PV was reported to contain phenolic acids (caffeic acid, chlorogenic acid, ellagic acid, ferulic acid, and protocatechuic acid) and flavonoids (rutin, quercetin, quercitrin, luteolin, and apigenin).3 These phenolic compounds have been extensively studied for their antioxidant and anti-inflammatory properties.4 The antioxidant effects of aqueous PV tea infused preparation (PVT) were previously reported.5 Limited data available to evaluate the effects of PVT on inflammation. More scientific evidence is required to examine the therapeutic benefits, with focus on antioxidant and anti-inflammatory activity, of the “cooling tea” brewed from this traditional herb.
Limited data is available to evaluate the antioxidant and anti-inflammatory properties of PVT. The study aimed to examine, using established in vivo markers of oxidative stress and inflammation, the antioxidant and anti-inflammatory actions of PVT in-vitro. The same study also evaluated the total flavonoid and phenolic contents, and total antioxidant capacities of PVT. The measured parameters of PVT were compared with those measured from green (GT) and black tea (BT).
Materials and Methods
Chemicals and materials
Arachidonic acid (AA), F2-isoprostanes (F2IP), F2-isoprostanes-d4, leukotriene B4 (LTB4), and leukotriene B4-d4 were purchased from Cayman Chemical (Ann Arbor, MI, USA). 1,1-diphenyl-2-picrylhydrazyl radical (DPPH), 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulfonate) (ABTS), 2,2′-azobis-(2-amidinopropane) hydrochloride (ABAP), 2,2′-azobis(2-methylpropionamidine) dihydrochloride (AAPH), 2,3,4,5,6-pentafluorophenylbromide, 12-myristate 13-acetate (PMA), 5ɑ-cholestane, acetic acid, ammonium ferrous sulfate, bis(trimethylsilyl) trifluoroacetamide, caffeic acid (CA), calcium ionophore, campesterol, catechin (C), fluorescein sodium salt, glutamine, guaiacol, hydrogens peroxide (50% by volume), isooctane, phosphate-buffered saline (PBS), potassium hydroxide, pyridine, R-phycoerythrin, phosphoric acid, quercetin (Q), rosmarinic acid (RA), sitosterol, sodium orthophosphate, disodium phosphate, [15N]-sodium nitrite, [15N]-sodium notrate, stigmasterol, streptomycin, superoxide dismutase (SOD), toluene, trichloroacetic acid (TCA), trolox, vitamin C, and xylenol orange were purchased from Sigma-Aldrich (St. Louis, MO, USA). Acetonitrile, ethyl acetate, methanol, ethanol, and sulfuric acid were purchased from Tedia (Fairfield, OH, USA). Ficoll-paque and dextran-500 were purchased from GE Healthcare (Uppsala, Sweden). The dried flowering tops of Prunella vulgaris, dried green tea leaves, and black tea leaves were used to prepare the tea. They were purchased from a renowned local herb shop, (Hockhua Tonic, Singapore).
Preparation of herbal tea
The PVT, GT, and BT were prepared by infusing the respective herbs or leaves (100 g dried mass) in deionised water (100 mL) at 80 °C for 15 minutes. The tea was filtered using filter paper (Whatman Filter Paper No. 1) before cooling to room temperature for subsequent extraction and analyses. Quintuplicates were performed for each tea (N=5).
Composition analysis
Extraction
Each of the PVT, GT, and BT (30 mL) was extracted using 100% methanol (10 mL x 3). The methanolic extract was adjusted to 30 mL using the rotary evaporator.
Total phenolic content
The total phenolic content in the extract was determined by the Folin-Ciocalteu’s assay.6 The total phenolic content was expressed as gallic acid equivalents per 100g.
Total and specific flavonoid content
The total flavonoid content was quantified by the modified Dowd colorimetric method.7 It was expressed as the quercetin-equivalent (QE) per 100g.
The total amounts of catechin and quercetin in the methanolic extract were measured using high-performance liquid chromatography (HPLC).6 Briefly, the extract was hydrolyzed in methanolic potassium hydroxide (3 mol/dm3) under nitrogen gas at 60 °C for 3 hours. The supernatant was filtered through a 0.45 µm cellulose acetate syringe filter before HPLC injection. The filtered supernatant (20 µL) was chromatographed isocratically on a reversed-phase C18 column (Merck Purospher® Star RP-18, 5 µm particle size, 8 A pore size, 100 × 4.60 mm) at a flow rate of 1.0 mL/min using a Waters HPLC series 2695 with mobile phase methanol, acetonitrile, and water (60:20:20, v/v/v). Catechin and quercetin were measured at 276 nm and 262 nm, respectively, and determined by comparing the catechin and quercetin integrated peak areas with respective, predetermined calibration curves.
Total plant sterol content
The amounts of total plant sterols in the PVT, GT, and BT were calculated as the sum of the amounts of the three specific PS (sitosterol, campesterol, and stigmasterol). The amounts of sitosterol, campesterol, and stigmasterol in PVT, GT, and BT were determined using Gas Chromatography-Mass Spectroscopy (GC-MS) as previously described.8 Briefly, internal standard (5ɑ-cholestane, 2 µmol) was added into the methanolic extract (20 mL). The mixture was hydrolyzed with ethanolic potassium hydroxide (final concentration 1 mol L-1) at 70 °C for 60 minutes in the dark. The lipids were extracted twice with hexane and ethanol, containing 12.5 mg/L BHT (20:1 v/v, 10 mL). The dried lipid extract was derivatized with pyridine-BSTFA with 1% TMCS (1:1 v/v, 200 µL) at 70 °C for 60 minutes. The derivatized extract was dried under nitrogen and reconstituted in isooctane (20 µL) before being injected into the GC-MS. The MS was operated in the electron ionization mode. The 5ɑ-cholestane, sitosterol, campesterol, and stigmasterol were monitored at m/z 357, 486, 472, and 484, respectively.
All analyses were performed in quintuplicate.
Antioxidant capacities
The antioxidant capacity of the methanolic extract was evaluated using the DPPH radical scavenging activity9, oxygen radical absorbance capacity (ORAC)10, total peroxyl trapping potential (TRAP)11, and trolox equivalent antioxidant capacity (TEAC)6 assays.
Briefly, for DPPH radical scavenging assay9, the methanolic extract (1 mL) was added into the freshly prepared ethanolic DPPH solution (48 mg L-1, 9 mL). The change in absorbance at 517 nm was measured. A calibration curve was prepared using vitamin C (0-500 mmol L-1). Absorbance was corrected for the solvent. The DPPH radical scavenging results were expressed in mmol L-1 vitamin C equivalents.
For ORAC assay10, the PV methanolic extract (20 µL) and fluorescein sodium salt (200 µL, 81.6 nmol L-1) were added into a well of a 96-well plate. Fluorescence (kexcitation = 485 nm, kemission = 528 nm) was measured. AAPH (75 µL, 200 mM) was then added to the well. Fluorescence was recorded every minute for over 40 minutes. A calibration curve was prepared with trolox standards (0-50 µmol L-1). Fluorescence was corrected for the solvent. The ORAC results were expressed as µmol L-1 trolox equivalents.
For TRAP assay11, the methanolic extract (1 mL) was incubated with TRAP reaction mixture (0.43 mg L-1 R-phycoerythrin and 0.3 g L-1 AAPH in 50 mmol L-1 PBS (pH 7.4), 2 mL) at 37 ºC for 5 minutes. Fluorescence (kexcitation = 540 nm, kemission = 565 nm) was measured in a quartz cuvette. A calibration curve was prepared with vitamin C (0 – 100 mmol L-1). Fluorescence was corrected for the solvent. The TRAP results were expressed as mmol L-1 vitamin C equivalents.
For TEAC assay9, the PV methanolic extract (40 µL) was mixed with the freshly prepared radical solution (20 mmol L-1 ABTS and 2.5 mmol L-1 ABAP stock solution in PBS, 1960 µL). The decrease in absorption at 734 nm was measured over 6 minutes. A calibration curve was prepared with trolox (0-1 mmol L-1). Absorbance values were corrected for the solvent. The TEAC results were expressed in mmol L-1 trolox equivalents. All experiments were performed at least in quintuplicate.
Cellular antioxidant and anti-inflammatory activities
The cellular experiments were performed with human peripheral blood neutrophils, which were freshly isolated from the neutrophil/erythrocyte pellet of fresh human whole blood after Ficoll-Paque gradient centrifugation and dextran sedimentation of red blood cells.12 The whole human blood was obtained in kind from the study researchers and as such did not require ethics approval. The freshly isolated neutrophils were resuspended in PBS at a concentration of 5 × 106 cells mL-1. Cell viability was assessed using trypan blue exclusion and was typically >98%.
The cellular antioxidant capacity of the metabolic extract of PVT was examined by measuring the inhibition of F2IP (stable marker of AA oxidation) and lipid hydroperoxides (LPO) formation in freshly isolated human blood neutrophils.7 The cellular anti-inflammatory of the same extract was evaluated by measuring the inhibition of LTB4 formation and myeloperoxidase (MPO) activity in freshly isolated human neutrophils.6,12 Briefly, the freshly isolated neutrophils (1 mL, 5 x 106 cells/mL PBS) were incubated with either PVT, GT, or BT methanolic extract (final concentration 1% v/v, 10 µL) and AA (final concentration, 10 µmol L-1) at 37 °C for 5 minutes before stimulation. To stimulate the F2IP formation, the neutrophils were incubated with PMA (final concentration, 200 nM) at 37 °C for 15 minutes. Cells treated with AA only and activated by PMA were used as positive controls, while cells treated with AA only without PMA activation were used as negative controls. For LPO productions, the neutrophils were stimulated with AAPH (final concentration, 5 mmol/L) at 37 °C for 15 minutes. Cells treated with AA only and activated by AAPH were used as positive controls, while cells treated with AA only without AAPH activation were used as negative controls. The neutrophils were incubated with calcium ionophore (final concentration, 200 nmol/L) at 37 °C for 15 minutes to stimulate the production of LTB4. Cells treated with AA only and activated by calcium ionophore were used as positive controls, while cells treated with AA only without calcium ionophore stimulation were used as negative controls. The supernatant from the neutrophil suspension was collected and stored at -80 °C before extraction and analysis of F2IP, LPO, and LTB4. F2IP and LTB4 were quantitated using stable isotope labeled GCMS.9,12 The formation of LPO was quantitated using the Ferrous Oxidation-Xylenol Orange assay.13 To assess the effects of PVT, GT, and BT on MPO activity, the treated neutrophils were resuspended in fresh PBS (1 mL) and lyzed by sonication. Untreated neutrophils were used as positive control. Functional MPO activity was determined by measuring its catalytic action on the oxidation of guaiacol in the presence of hydrogen peroxide.13
The antioxidant enzyme activity of the treated human neutrophils was assessed by determining the enzyme activities of SOD and peroxidases. The neutrophil pellet obtained after the designated treatment (PVT, GT, BT, CA 10 µmol L-1, RA 10 µmol L-1, Q 10 µmol L-1, and C 10 µmol L-1) was lyzed by sonication for 5 minutes at 4 ºC. The SOD activity in the lyzed cell supernatant was determined using a commercial SOD colorimetric activity assay kit (Thermo Fisher Scientific, Waltham, MA, USA). The peroxidase activity in the lyzed cells was measured using a commercial peroxidase activity assay kit (Merck, Darmstadt, GERMANY). The SOD and peroxidase enzyme activities were expressed as the percentage of activity relative to the untreated positive control.
The effects of the three tea treatments on the NOS activity were determined by measuring the amounts of nitrite and nitrate formed by the human neutrophils after their respective treatments.14 Briefly, the freshly isolated human blood neutrophils (5×106 cells/mL in PBS, 1 mL) was incubated with either PVT, GT or BT methanolic extract (final concentration 1% v/v, 10 μL) and arginine (final concentration, 10 μmol/L) at 37 °C for 5 minutes prior to stimulation. The neutrophils were stimulated with PMA (final concentration, 200 nmol L-1) at 37 ºC for 5 minutes. Untreated cells with arginine were used as positive controls while untreated cells without arginine served as negative controls. The cell supernatant was collected at the end of the incubation and stored at -80 ºC before nitrite and nitrate analysis. The releases of nitrite and nitrate from stimulated neutrophils were measured by stable isotope labeled GC-MS as previously described.14
Statistical analyses
Statistical analyses were performed using Microsoft Excel (version 2103, April 2021). Data (N=5 for each tea) were assumed to be normally distributed and presented as mean ± standard deviation (SD). Differences between one group and a predefined value were compared using one-sample t-tests. Differences between two or more than two groups were compared using two-sample independent t-test and ANOVA with the Bonferroni post-hoc test, respectively. A significant difference or correlation was observed when p<0.05.
Results
Phytochemical analysis
The total phenolic content of PVT (137±17 mg GAE/ 100g PVT) was significantly greater than that of GT (77±17 mg GAE/ 100g GT), but did not differ from that of BT (121±9 mg GAE/ 100g BT). The PVT (24±7 mg QE/ 100g PVT) contained significantly higher flavonoid content than GT (11±3 mg QE/ 100g GT) and BT (11±2 mg QE/ 100g BT). GT and BT did not differ in total flavonoid contents. PVT contained significantly higher total quercetin content (2.58±0.32 mg/ 100g PVT) than GT (1.74±0.06 mg/ 100g GT) and BT (1.62±0.13 mg/ 100g BT) and significantly lower total catechin content (51.2±5.5 mg/ 100g PVT) than GT (62.0±6.0 mg/ 100g GT). GT had significantly higher total catechin content than BT (48.4±6.3 mg/ 100g BT), but the two teas did not differ in total quercetin contents. PVT (3.14±1.04 ppm), GT (2.95±0.90 ppm), and BT (3.33±1.14 ppm) contained significant amounts of total plant sterols. The total plant sterols amounts did not differ between the three teas.
Antioxidant capacities
PVT showed significantly stronger DPPH radical scavenging activity than GT and BT (Figure 1a). GT and BT did not differ in their DPPH radical scavenging activities (Figure 1a). The three teas, PVT, GT, and BT, did not differ in their ORAC and TRAP activities (Figures 2b and 2c). PVT demonstrated significantly stronger TEAC activity than GT and BT (Figure 1d). Significant difference in TEAC activities was also observed between GT and BT (Figure 1d).
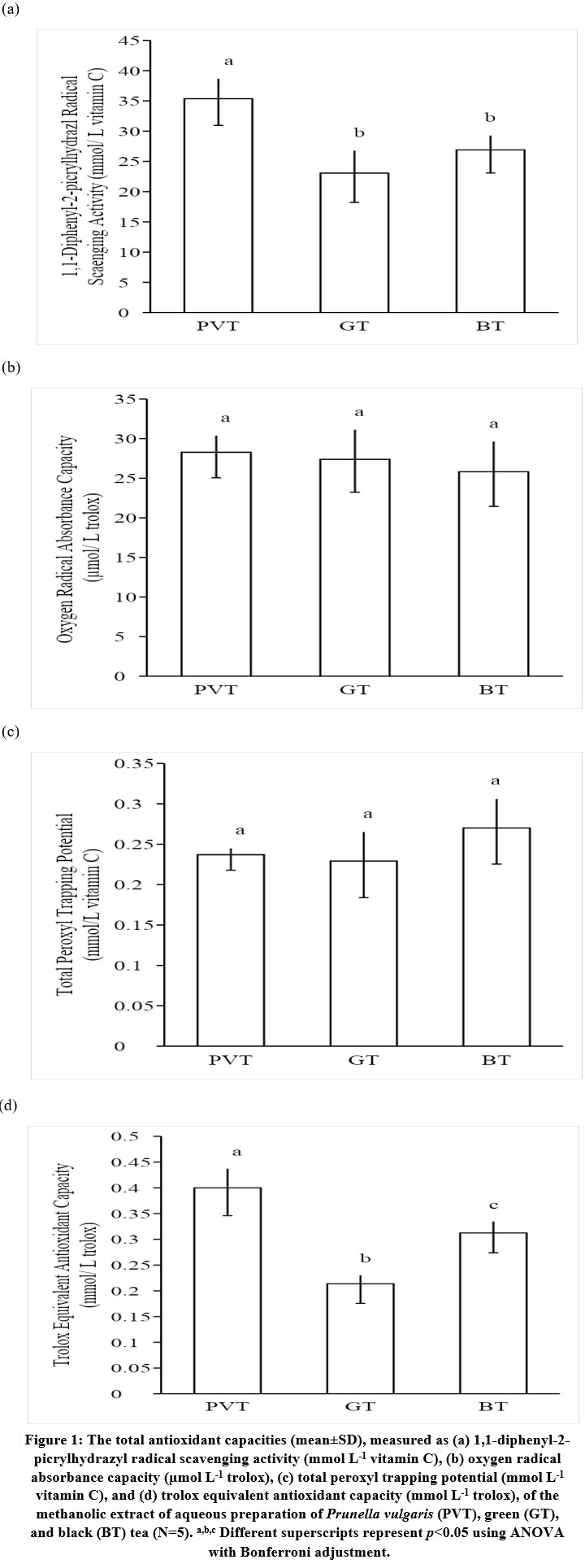 |
Figure 1: The total antioxidant capacities (mean ± SD), measured as (a) 1,1-diphenyl-2-picrylhydrazyl radical scavenging activity (mmol L-1 vitamin C), (b) oxygen radical |
Cellular experiments
PVT, GT, and BT significantly decreased neutrophil F2IP formation (Table 1). The amounts of F2IP produced by human neutrophils were significantly lower when the cells were incubated with PVT when compared with GT and BT (Table 1). Significant difference in cellular F2IP formation was absent between GT and BT (Table 1). PVT inhibited LPO formation in human neutrophils to a significantly greater extent than GT and BT (Table 1). Significant difference in cellular LPO formation was observed between GT and BT (Table 1). The LPO formations were significantly reduced by the PVT, GT, and BT treatments (Table 1). The SOD activity of freshly isolated human neutrophils was significantly increased by PVT, but not by GT and BT (Figure 2a). All three treatments significantly elevated cellular peroxidase activities (Figure 2b). PVT increased SOD and PO activities significantly more than GT and BT (Figures 2a and 2b). The difference between the effects of GT and BT on SOD and PO activities did not attain significance (Figures 2a and 2b). The SOD and PO enzyme activities were significantly elevated when the neutrophils were treated with Q or C (Figures 2a and 2b). Significant effects were absent with CA and RA (Figures 2a and 2b).
PVT, GT, and BT treatments significantly inhibited the LTB4 production of the human neutrophils (Table 1). Human neutrophils treated with PVT produced significantly lower amounts of LTB4 than those treated with GT and BT (Table 1). BT inhibited cellular LTB4 formation to a significantly greater extent than GT (Table 1). All three tea treatments significantly reduced the MPO activity of the human neutrophils (Table 1). The MPO activity of the human neutrophils was inhibited by PVT to a significantly greater extent than by GT and BT (Table 1). GT and BT did not differ in their MPO inhibitory activities (Table 1). The nitrite and nitrate productions by the stimulated neutrophils were significantly augmented by PVT (nitrite 130.2±6.8%, nitrate 134.2±3.8%) and GT (nitrite 112.3±6.8%, nitrate 105.0±4.2%). BT (nitrite 99.5±5.5%, nitrate 95.9±8.6%) did not significantly affect the nitrite and nitrate production. Significant differences were observed between the three treatments.
Table 1: (a) F2-isoprostanes, (b) lipid hydroperoxides, (c) leukotriene B4 formations, and (d) myeloperoxidase activity (% relative to positive controls, mean±SD) from freshly isolated human blood neutrophils after incubating with the methanolic extracts of aqueous Prunella vulgari tea (PVT), green tea (GT), and black tea (BT) (N=5).
| PVT | GT | BT | |
| (a) F2-isoprostanes (% relative to positive control) | 45.3±6.3*,a | 75.8±6.3*,b | 77.5±8.3*,b |
| (b) Lipid hydroperoxides (% relative to positive control) | 80.5±8.1*,a | 64.0±5.7*,b | 71.6±3.6*,c |
| (c) Leukotriene B4 (% relative to positive control) | 51.9±3.1*,a | 82.6±3.6*,b | 72.1±4.3*,c |
| (d) Myeloperoxidase activity (% relative to positive control) | 81.4±3.9*,a | 92.3±3.2*,b | 90.5±3.0*,b |
| * p<0.05 vs. 100% using one-sample t-tests.a,b,c Different superscripts represent p<0.05 using ANOVA with Bonferroni adjustment. | |||
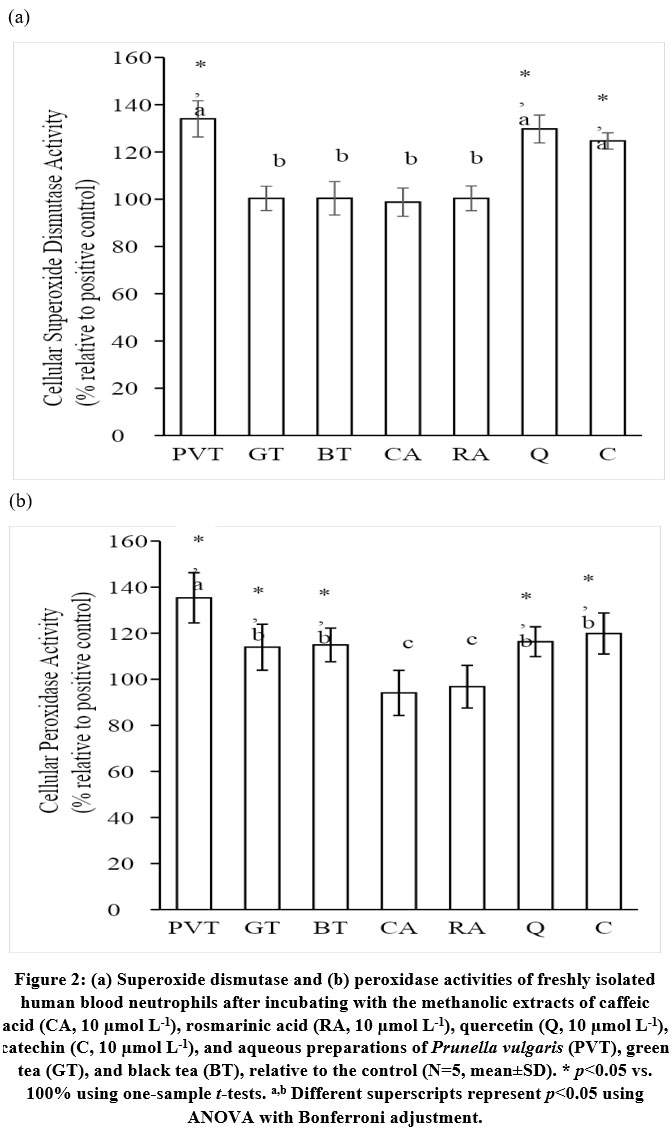 |
Figure 2: (a) Superoxide dismutase and (b) peroxidase activities of freshly isolated |
Discussion
The total phenolic, flavonoid, and quercetin contents of PVT were measured to be significantly higher than those of GT and BT. The three teas contained similar amounts of total plant sterols. These phytochemical compounds have been extensively studied as antioxidants and anti-inflammatory agents.4 The health-benefiting properties of green and black tea consumptions have been chiefly attributed to their phenolic and flavonoid contents.15,16 The presence of significant amounts of these compounds in PVT is likely to confer similar physiological effects. Consequently, PVT would most likely exert antioxidant and anti-inflammatory activities.
Antioxidants exert their activities via various direct or indirect pathways.17 One of the most studied antioxidant pathways is the radical scavenging action.17 The results from the DPPH, ORAC, TRAP, and TEAC radical scavenging assays suggested that PVT, GT, and BT exhibited significant antioxidant capacity via the radical scavenging mechanisms, and PVT demonstrated stronger such action than GT and BT. Aqueous extract of PV demonstrated strong superoxide- and hydroxyl radical-scavenging activity but exerted only a slight pro-oxidant effect.5 Methanolic extract of PV (25.7% rosmarinic acid) demonstrated stronger DPPH radical scavenging activity than caffeic acid and rosmarinic acid.3 The same organic extract also inhibited copper-mediated low-density lipoprotein oxidation to greater extents than caffeic acid and rosmarinic acid, inferring that PV contains more potent antioxidants than rosmarinic acid and caffeic acid.3 As shown earlier, PV contained significant amounts of flavonoids, like catechin and quercetin, which can further contribute to its antioxidant capacity. Mechanistically, radical scavenging occurs through hydrogen atom transfer (HAT) and/ or electron transfer (ET). The four radical scavenging assays used in the experiments differ in their reaction mechanisms and can be employed to evaluate the radical scavenging mechanisms.17 ORAC and TRAP measure the radical scavenging capacity via the HAT mechanism, as the reactions involve the transfer of hydrogen atoms from the probe to the radical.17 The reactions in DPPH and TEAC assays, on the other hand, involve the transfer of electrons, not hydrogen atoms, between the probe and radicals and thereby determine the radical scavenging activity via the ET mechanism.17 PVT demonstrated significant radical scavenging capacities via both HAT and ET, but showed stronger reactivities via ET when compared with GT and BT. Flavonoids, like quercetin and catechin, scavenge radicals predominantly via the ET mechanism.18,19 Phenolics, on the other hand, neutralize the radicals predominantly via the HAT mechanism.20 GT and BT had been shown to exhibit significant radical scavenging activities. The flavonoids and phenolics in PVT may contribute to the strong radical scavenging property and its associated mechanisms. The comparative stronger radical scavenging activity may also be accounted for by their higher contents. F2IP is usually formed by the oxidation of AA via the radical chain mechanism, and it is an established in vivo marker of oxidative stress.21 LPO is the radical-propagated oxidation product of cellular lipids (usually from the cell membrane).22 The reductions in F2IP and LPO formation in vitro provide physiological evidence on the antioxidant property of PVT via the radical scavenging activity. PVT reduced F2IP and LPO formation to a stronger extent than GT and BT. The cellular cell experiment result is consistent with the radical scavenging capacity of PVT relative to those of GT and BT. In addition to direct radical scavenging activity, PVT also significantly elevated the antioxidant enzyme, SOD and PO, activities. The results of the separate experiments with CA, RA, Q, and C suggested that augmentation of antioxidant enzyme activity may be attributed to the PVT flavonoid contents instead of its phenolic composition. The elevations of SOD and PO activity were previously reported for quercetin and catechin, but not for simpler phenolic molecules.23,24
LTB4 is a pro-inflammatory chemokine produced by blood neutrophils and macrophages at the site of inflammation, and is usually measured as the marker of local inflammation.25 Flavonoids, quercetin and catechin, reduced LTB4 formation dose-dependently in vitro and in vivo, possibly via inhibition of 5-lipoxygenase.26 Similar actions were observed with phenolic compounds.27 All three studied teas also contained significant amounts of total plant sterols, which had been shown to inhibit 5-lipoxygenase activity.8 It is not surprising that the flavonoid-, phenolic-, and plant sterol-rich PVT, GT, and BT demonstrated strong LTB4 formation inhibitory action. MPO is a type of heme peroxidases mainly expressed in neutrophils and monocytes.28 It is a critical component of the host immune system producing reactive species combating pathogens.29 Elevated MPO levels and MPO-derived reactive species are also found to be associated with inflammation and increased oxidative stress.28 Inhibiting MPO activity without compromising pathogen killing by the innate immune system has become an important direction to reduce MPO-associated host tissue damage.29 MPO activity and MPO-derived products can be diminished by selected flavonoids, such as quercetin30 and catechin31, and phenolic compounds, like caffeic acid and chlorogenic acid.32 The MPO inhibitory actions of PVT, GT, and BT emulate those of the flavonoids and phenolic compounds. The results suggested that PVT treatment significantly increased the inducible nitric oxide synthase (iNOS) activity of the freshly isolated neutrophils. The iNOS in human blood neutrophils have been reported to modulate the immune responses of the neutrophils and the associated inflammatory events.33 Previous studies provided evidence to support the anti-inflammatory properties of PVT. PV total flavonoids inhibited the growth of tumors in H22 tumor-bearing mice by promoting apoptosis in liver cancer cells via upregulating expressions of pro-inflammatory interleukin-6, tumor necrosis factor-α (TNFɑ), and IFN-γ.34 Using ligand-based pharmacophore generation approach and virtual screening against natural products libraries, PV contains compounds that target pro-inflammatory mediators TNFα .35 PV aqueous extract increased the mRNA and protein expressions and the activity of endothelial nitric oxide synthase in human umbilical vein endothelial cells (HUVEC).36 The results from this study illustrated the anti-inflammatory effects of PVT via 5-lipoxygenase, MPO, and iNOS pathways.
It is important to understand the potential physiological effects of herbs through scientific and clinical evaluation in the most physiological relevant manner. The phenolic contents were directly determined from the herb in previous studies. In this study, the phenolics were extracted and determined from an aqueous herbal preparation, which is the actual delivery form to the consumers. The phenolic content represents the actual exposure to the consumers. All the cellular experiments performed in the study were carried out using freshly isolated human blood neutrophils, which is a physiological-relevant oxidative stress and inflammation cell model.37 The results from these experiments may be more relevant and translatable to the actual in vivo conditions. The cellular antioxidant and anti-inflammatory activities were examined by measuring the established in vivo markers of oxidative stress (F2IP and LPO) and inflammation (LTB4 and MPO activity). The results were obtained mainly from cellular works, which may not represent the activities in vivo. Well-designed clinical studies are required to ascertain these cellular observations.
PVT exhibited significant antioxidant activities via ET radical scavenging and upregulating antioxidant enzymes. It also exerted anti-inflammatory effects through the modulations of pro-inflammatory 5-lipoxygenase, MPO, and iNOS pathways. The measured antioxidant and anti-inflammatory effects may be attributed to PV’s phenolic and flavonoid contents, and were greater than those from GT and BT. These effects may explain in part the reported therapeutic properties of PVT.
Acknowledgment
Nil.
Conflict of Interest
The author(s) declares no conflict of interest.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
- Marchio P , Guerra-Ojeda S., Guerra-Ojeda,, J. M., Aldasoro M. V., Victor M., Mauricio M. D. Targeting early atherosclerosis: A focus on oxidative stress and inflammation. Oxidat. Med. Cell Longev. 2019; 2019. https://doi.org/10.1155/2019/8563845
CrossRef - Devrim-Lanpir A., Bilgic P., Kocahan T., Deliceoğlu G., Rosemann T., Knechtle B. Total dietary antioxidant intake including polyphenol content: Is it capable to fight against increased oxidants within the body of ultra-endurance athletes? Nutrients. 2020;12:1877-1889. https://www.mdpi.com/2072-6643/12/6/1877
CrossRef - Wang S., Wang X., Dai Y, Ma M., Rahman K., Nian H., Zhang H. Prunella vulgaris: A comprehensive review of chemical constituents, pharmacological effects and clinical applications. Pharmaceut. Des. 2019; 25:359-369. http://researchonline.ljmu.ac.uk/id/eprint/10383/
CrossRef - Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. Food Biochem. 2020;44:e13394-e13405. https://doi.org/10.1111/jfbc.13394
CrossRef - Liu F., Ng T. B. Antioxidative and free radical scavenging activities of selected medicinal herbs. Life Sci. 2000;66:725-735. https://doi.org/10.1016/S0024-3205(99)00643-8
CrossRef - Loong C, Leo L., Loke W. M. Sugar, vitamin C, and polyphenols in commercial apple beverages and their effects on antioxidant and anti-inflammatory activities in vitro. J. Food Bioact. 2021;13:52-61. https://doi.org/10.31665/JFB.2020.13259
CrossRef - Ling L., Loong C, Loke W. M. Food additives in commercial cocoa beverage products and their effects on total polyphenol contents, cellular antioxidant and anti-inflammatory activities. Curr. Res. Nutr. Food Sci. 2021;9:20-30. http://dx.doi.org/10.12944/CRNFSJ.9.1.03
CrossRef - Ho X. L., Liu J. J. H., Loke W M. Plant sterol-enriched soy milk consumption modulates 5-lipoxygenase, 12-lipoxygenase and myeloperoxidase activities in healthy adults – A randomized controlled trial. Free Rad. Res. 2016;50:1396-1407. http://dx.doi.org/10.1080/10715762.2016.1252839
CrossRef - Leung H. H., Yau Y. F., Leung K. S., Lee Y. Y., Oger C., Durand T., Galano J. M., Loke W. M., Lee J. C. Garlic supplementation modified enzymatic omega-6 polyunsaturated fatty acid oxidation in mild hypercholesterolemia. J. Lipid Sci. Technol. 2019;121:1900069-1900080. https://onlinelibrary.wiley.com/doi/abs/10.1002/ejlt.201900069
CrossRef - Ruefer C. E., Kulling S. E. Antioxidant activity of isoflavones and their major metabolites using different in vitro J. Agri. Food Chem. 2006;54:2926-2931. https://doi.org/10.1021/jf053112o
CrossRef - Ghiselli A., Serafini M., Maiani G., Azzini E., Ferro-Luzzi A. A fluorescence-based method for measuring total plasma antioxidant capability. Free Radical Biol. Med. 1995;18:29-36. https://doi.org/10.1016/0891-5849(94)00102-P
CrossRef - Tsen S. Y,, Tan X. Y., Tan Y. M., Yan B. Y., Loke W. M. Relative inhibitions of 5-lipoxygenase and myeloperoxidase and free-radical scavenging activities of daidzein, dihydrodaidzein, and equol. Med. Food. 2016;19:543-548. https://doi.org/10.1089/jmf.2015.3557
CrossRef - HoX L., Loke W. M. Combination of specific probiotic strains and dietary polyphenols exhibit synergistic cellular antioxidant and anti-inflammatory potentials. Int. J. Probio. Prebio. 2021;16:31-38. https://doi.org/10.37290/ijpp2641-7197.16:31-38
CrossRef - HoX L., Loke W. M. Dietary plant sterols supplementation increases in vivo nitrite and nitrate production in healthy adults: A randomized, controlled study. J. Food Sci. 2017;82:1750-1756. https://doi.org/10.1111/1750-3841.13752
CrossRef - Jakubczyk K., Kochman J., Kwiatkowska A., Kałduńska J, Dec K., Kawczuga D., Janda, K. Antioxidant Properties and nutritional composition of matcha green tea. Foods. 2020;9:483-487. https://www.mdpi.com/2304-8158/9/4/483
CrossRef - Zhang H, Qi R., Mine Y. The impact of oolong and black tea polyphenols on human health. Food Biosci. 2019;29:55-61. https://doi.org/10.1016/j.fbio.2019.03.009
CrossRef - Huang D, Ou B., Prior R. L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005;53:1841-1856. https://doi.org/10.1021/jf030723c
CrossRef - Amorati R., Baschieri A., Cowden A., Valgimigli L. The Antioxidant Activity Of Quercetin In Water Solution. Biomimetics. 2017;2:9-22. https://doi.org/10.3390/biomimetics2030009
CrossRef - Yang Y., Zeng H., Zhang Q., Bai X., Liu C., Zhang, Y. H. Direct electron transfer and sensing performance for catechin of nano-gold particles-polymer nano-composite with immobilized Laccase. Phys. Lett. 2016;658:259-269. https://doi.org/10.1016/j.cplett.2016.06.063
CrossRef - Bietti M., et al. 2019. Evaluation of polar effects in hydrogen atom transfer reactions from activated phenols. Org. Chem. 84:1778-1786.
CrossRef - Bietti M., Cucinotta E., DiLabio G. A., Lanzalunga O., Lapi A., Mazzonna M., Romero-Montalvo E., Salamone M. Urinary 8-isoprostane as a biomarker for oxidative stress: A systematic review and meta-analysis. Lett. 2019;328:19-27. https://doi.org/10.1021/acs.joc.8b02571
CrossRef - Shi F., Ma C., Ji C., Li M., Liu X. Han Y. Serum lipid oxidative stress products as risk factors are the candidate predictive biomarkers for human abdominal aortic aneurysms. Appl. Thromb. Hemost. 2020;26:1-12.
CrossRef - Robaszkiewicz A, Balcerczyk A., Bartosz G. Antioxidative and prooxidative effects of quercetin on A549 cells. Cell Biol. Int. 2007;31:1245-1250. https://doi.org/10.1016/j.cellbi.2007.04.009
CrossRef - Simos Y. V., Verginadis J. I., Toliopoulos I. K., Velalopoulou A. P., Karagounis I. V., Karkabounas S. C., Evangelou A. M.. Effects of catechin and epicatechin on superoxide dismutase and glutathione peroxidase activity, in vivo. Redox Rep. 2012;17:181-186. https://doi.org/10.1179/1351000212Y.0000000020
CrossRef - Horii Y., Nakaya M., Ohara H., Nishihara H., Watari K., Nagasaka A., Nakaya, T., Sugiura Y., Okuno T., Koga T., Tanaka A., Yokomizo T., Kurose H. Leukotriene B4 receptor 1 exacerbates inflammation following myocardial infarction. FASEB J. 2020;34:8749-8763. https://doi.org/10.1096/fj.202000041R
CrossRef - Loke W. M., Proudfoot J. M., Hodgson J. M., McKinley A. J., Hime N., Magat M., Stocker R., Croft K. D.. Specific dietary polyphenols attenuate atherosclerosis in Apolipoprotein E-knockout mice by alleviating inflammation and endothelial dysfunction. Thromb. Vasc. Biol. 2010;30:749-757. https://doi.org/10.1161/ATVBAHA.109.199687. ATVB_2010 2010
CrossRef - Giménez-Bastida J. A., González-Sarrías A., Laparra-Llopis J., Moisés J., Schneider C., Espín, J. C. Targeting mammalian 5-lipoxygenase by dietary phenolics as an anti-inflammatory mechanism: A systematic review. J. Molec. Sci. 2021;22:7937-7945. https://www.mdpi.com/1422-0067/22/15/7937
CrossRef - Ndrepepa G. Myeloperoxidase – A bridge linking inflammation and oxidative stress with cardiovascular disease. Chim. Act. 2019;493:36-51. https://doi.org/10.1016/j.cca.2019.02.022
CrossRef - Davies M.,, Hawkins C. L. The role of myeloperoxidase in biomolecule modification, chronic inflammation, and disease. Antioxid. Redox Signal. 2020;32:957-981. https://doi.org/10.1089/ars.2020.8030
CrossRef - Tian R., Jin Z., Zhou L, Zeng X., Lu N. Quercetin attenuated myeloperoxidase-dependent HOCl generation and endothelial dysfunction in diabetic vasculature. Agric. Food Chem. 2021;69:404-413. https://doi.org/10.1021/acs.jafc.0c06335
CrossRef - Sunil M. A., Sunitha V. S., Santhakumaran P., Mohan M.C., Midhun Sebastian J., Radhakrishnan E. K., Mathew J. Protective effect of (+)–catechin against lipopolysaccharide-induced inflammatory response in RAW 264.7 cells through downregulation of NF-κB and p38 MAPK. Inflammopharmacology. 2021;29:1139-1155. https://doi.org/10.1007/s10787-021-00827-6
CrossRef - Kolgazi M., Cilingir S., Yilmaz O., Gemici M., Yazar H., Ozer S., Acikel-Elmas M., Arbak S., Suyen G. G. Caffeic acid attenuates gastric mucosal damage induced by ethanol in rats via nitric oxide modulation. Biol. Interact. 2021;334:109351-109356. https://doi.org/10.1016/j.cbi.2020.109351
CrossRef - Saini R and Singh S. Inducible nitric oxide synthase: An asset to neutrophils. J. Leukocyte Biol. 2019;105:49-61. https://doi.org/10.1002/JLB.4RU0418-161R
CrossRef - Song Y., Kang L., Tian S., Cui, L., Li Y., Bai M., Fang X., Cao L., Coleman K., Miao M. Ethnopharmacol. 2021;273:113891-113903. https://doi.org/10.1016/j.jep.2021.113891
CrossRef - Zaka M., Sehgal S., Arslan S., Abbasi B. H.. Comparative in silico analyses of Cannabis sativa, Prunella vulgaris and Withania somnifera compounds elucidating the medicinal properties against rheumatoid arthritis. Mol. Graph. Model. 2017;74:296-304. https://doi.org/10.1016/j.jmgm.2017.04.013
CrossRef - Xia N., Bollinger L., Steinkamp-Fenske K., Förstermann U., Li H. Prunella vulgaris L. upregulates eNOS expression in human endothelial cells. J. Chinese Med. 2010;38:599-611. https://doi.org/10.1142/s0192415x10008081
CrossRef - Silvestre-Roig C., Braster Q., Ortega-Gomez A., Soehnlein O. Neutrophils as regulators of cardiovascular inflammation. Rev. Cardiol. 2020;17:327-340. https://doi.org/10.1038/s41569-019-0326-7
CrossRef
Accepted on: 14 Aug 2022
Second Review by: Husniye Kayalar Turkey
Final Approval by: Dr. Dariusz Dziki





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








