
Introduction
NCDs are long-term disease conditions that develop gradually due to genetic, physiological, environmental, lifestyle, and dietary factors. The most prevalent NCDs globally include diabetes, cancer, chronic respiratory diseases, and cardiovascular disorders. The incidence rate of NCDs is increasing globally and nationally. 1 Cancer, marked by uncontrolled cell proliferation and potential metastasis, is a major global health threat. 2 Every day, cancer is detected in 52,900 people, causing more than 27,000 deaths. Projections suggest that by 2040, the global burden of cancer will reach 28 million new cases and 16.2 million deaths annually.3 Conventional cancer treatments like surgery, chemotherapy, and radiotherapy often come with significant adverse effects. However, recent advancements in oncology have introduced innovative approaches such as ablation therapy, catalytic cancer therapy, iron-dependent cell death therapy, gene modification therapy, nanoparticle-based therapy, radionics, stem cell therapy, sonodynamic therapy, and targeted therapy. While these advanced therapies offer promising alternatives for cancer treatment, their high costs create significant barriers to accessibility, making them unaffordable for individuals from various socio-economic backgrounds. 4
Findings from medical and nutrition research indicate that dietary agents enhance anticancer activity and regulate several physiological functions. In the field of cancer research and therapy, scientists are currently exploring numerous chemical components produced from plants that can effectively fight and prevent cancer. 5 Phytochemicals/phytoconstituents/ non-nutritive compounds present in herbs, leaves, vegetables, fruits and other plant sources have attracted considerable attention because of their prospective roles in cancer inhibition and therapy due to the complexity of their structures, their unique biological effects, affordability, ease of use, and comparatively fewer harmful side effects. 6,7 The anticarcinogenic properties of these non-nutritive compounds include modulation of molecular signaling pathways, inhibition of early-stage carcinogenesis, protein modifications, and interaction with specific molecular signals. 8
Good nutrition is vital for maintaining metabolic homeostasis. Inclusion of specific foods and beverages in the diet can alter metabolic pathways by enhancing or diminishing them. Diets mainly comprised of plants include a broad variety of consumption habits marked by reduced animal-derived goods and a predominance of plant-based foods abundant in various phytochemical groups. 9 Herbs, spices, legumes, whole grains, vegetables, fruits, oilseeds, and nuts provide essential micronutrients, including vitamins, minerals, enzymes, and phytochemicals, all contributing to cellular protection and cancer prevention. Epidemiological and mechanistic studies suggest that adherence to a plant-based diet that is well balanced, with limited consumption of red meat and meats that are processed meats, is linked with a lower incidence of breast, prostate, colorectal, and lung cancers. 10, 11 Regarding diet patterns for the prevention of cancer, prioritizing suitable plant-centred food selections is essential. This review systematically compiles and evaluates the phytochemicals and bioactive compounds present in various plant-based food groups, delineating their molecular mechanisms of cancer prevention as evidenced in preclinical models and human clinical studies.
Materials and Methods
This review compiles and analyzes the current scientific evidence on the anticancer potential of phytochemicals, emphasizing their molecular mechanisms and translational relevance from preclinical models to clinical applications. A literature survey was performed in various research databases such as Google Scholar, PubMed, Scopus, and Web of Science, to identify pertinent peer-reviewed publications. The search included original research articles and review papers published between 2018 and 2025. The following keywords and Boolean operators were used to refine the search:
(“phytochemicals” OR “plant-derived compounds”) AND (“anticancer activity” OR “cancer therapy”)
(“natural compounds” AND “molecular mechanisms” AND “cancer”)
(“preclinical studies” OR “in vitro” OR “in vivo“) AND (“phytochemicals” AND “oncogenic pathways”)
(“clinical trials” AND “phytochemicals” AND “cancer treatment”)
The manuscripts for this review article were chosen upon fulfilling certain eligibility and exclusion criteria.
Inclusion criteria
Studies investigating the anticancer effects of phytochemicals with mechanistic insights.
Research papers detailing apoptotic pathways, autophagy modulation, angiogenesis inhibition, cell cycle regulation, or interactions with other oncogenic pathways.
Preclinical studies (both in vitro and in vivo) and clinical trials evaluating phytochemical-based cancer therapies.
Peer-reviewed publications in English with clearly defined methodologies and experimental outcomes.
Exclusion criteria
Studies lacking mechanistic insights or focusing solely on general phytochemical properties without cancer-specific findings.
Non-peer-reviewed sources, conference abstracts, and unpublished reports.
Phytochemicals as targeted cancer therapeutic agents
In recent years, growing scientific evidence has highlighted the powerful role of phytochemicals in reducing cancer risk. Phytochemicals present in plant-based foods exert protective effects against cancer through multiple molecular mechanisms. Naturally occurring compounds, including flavonoids, phytosterols, phenolic acids, carotenoids, and stilbenes, help combat cancer by inducing apoptosis, inhibiting angiogenesis, and modulating inflammatory and oxidative stress pathways. 12
Flavonoids
Flavonoids are natural compounds in plants, consisting of 2 rings of benzene connected by 3 carbon atoms, creating a C6-C3-C6 structure. They are classified into subclasses, including chalcones, dihydrochalcones, flavones, and flavanols, based on the oxidation level of the central chain and the B-ring attachment. 13 Common dietary sources of flavonoids include EGCG (cocoa and green tea), hesperidin and hesperetin (citrus fruits), quercetin in (apples, berries, broccoli, grapes, red onions, and red wine), luteolin (carrots, celery, chili peppers, lettuce, and spinach), and apigenin (artichokes, chamomile, oregano, and parsley). 14 Flavonoids reduce tumor progression through apoptosis and cell-cycle regulation. Flavonoids cause G2/M phase arrest and promote cell death in breast cancer cells by altering two pathways (p53 and PTEN). 15 EGCG regulates the 67-KDa laminin receptor, influences JAK/STAT, MAPK, and PI3K/AKT signaling pathways, and affects estrogen and androgen receptors in breast and prostate tumors, respectively. 16 Quercetin induces DNA damage, decreases BCL2 levels, and activates caspase-3 to trigger apoptosis. It sensitizes prostate cancer cells to paclitaxel by modulating proteins like GRP78 and hnRNPA1. 17 Apigenin’s anti-angiogenic properties are associated with cell cycle cessation, apoptosis induction, and regulation of signaling pathways. 18
Phytosterols
Phytosterols are plant-derived compounds resembling cholesterol, differing mainly by an extra hydrocarbon chain at the C-24 site. Over 250 phytosterols have been identified, with campesterol, β-sitosterol, stigmasterol, β-sitostanol, and campestanol being the most common in food sources. 19 These compounds show anticancer properties by enhancing pro-apoptotic signals that cause cell death by increasing calcium concentration in the cytosol and mitochondrial region. Phytosterols promote the overexpression of unfolded protein response signals and endoplasmic reticulum-mitochondria axis signals, leading to endoplasmic reticulum stress and cancer cell death. They also decrease the production of ROS, facilitating cell death via intrinsic mitochondrial apoptotic pathways or extrinsic death receptor pathways. Phytosterols hinder cell cycle progression by suppressing PCNA expression and disrupting the MAPK pathway, while also reducing cell migration and aggregation. 20 They help the immune system identify and kill cancer-causing cells. 21 β-sitosterol inhibits the propagation of human colon cancer cells by obstructing the LEF-1-mediated Wnt/β-catenin pathway. The increased expression of LEF-1 in intestinal cancer triggers this pathway, causing the growth of tumors. 22 β-Sitosterol impedes the advancement and invasiveness of HCT116 CC cells by lowering LEF-1 levels, disrupting the pathway, and diminishing key downstream targets, including Survivin and CCND1, supporting its effectiveness as a cancer-fighting agent.
Phenolic acids
The cancer-fighting ability of phenolic acids comes from their antioxidant effects. They act as free radical scavengers and metal binding agents, strengthen the body’s antioxidant defense mechanisms, and regulate key proteins and transcription factors (NRF2). 23, 24 Their role involves blocking cell growth via the ERK1/2 pathway, D-type cyclins, and CDK, alongside hindering angiogenesis by targeting VEGF and MIC-1, suppressing oncogenic signaling (PI3K/AKT), inducing apoptosis, and inhibiting cellular migration and metastasis. Their ability to fight against cancer is amplified through epigenetic influencers and resistance regulators, effectively reducing cancer cell division, inhibiting metastasis, and promoting cell death. 25 Phenolic acids, such as caffeic, dihydro cinnamic, and p-coumaric acids extracted from propolis, along with EGCG extracted from green tea, act as epigenetic agents for TNBC. 26 Administration of coumaric acid and EGCG led to a marked decline in cell viability across four TNBC cell lines (BT-20, BT-549, MDA-MB-231, and MDA-MB-436). Molecular docking predicted that these compounds bind to the MTAse territory of human DNMT1, competing with its endogenous inhibitor SAH. While EEP did not modify overall DNA methylation, EEP and EGCG were found to cause demethylation of the RASSF1A gene in BT-549 cells. EEP treatment also influenced RASSF1 protein expression, highlighting propolis as a potential agent for epigenetic therapies aimed at DNA methylation modulation. Chlorogenic acids present in tea help mitigate disease risk by regulating signaling pathways involved in inflammation and oxidative damage, regulating essential key cellular functions such as proliferation, apoptosis, and immune responses. By interacting with specific molecular mechanisms, chlorogenic acids suppress tumor growth. Research studies of phenolic acids in ovarian cancer have identified promising anticancer effects. GA suppresses tumor development via the PTEN/AKT/HIF-1α pathway. Salicylic acid regulates multiple pathways, including Wnt/β-catenin, and targets ovarian cancer stem cells. EA prevents angiogenesis and cisplatin resistance, facilitating apoptosis. PCA reduces cancer cell growth by inducing apoptosis and regulating oxidative stress. CA enhances cisplatin cytotoxicity, while coumaric acid and ferulic acid act as adjuvants, modulating oxidative stress and signaling pathways. 27
Stilbenes
Dietary stilbenes are polyphenolic compounds found in many plants, such as berries, grapes, and medicinal herbs. Certain examples of notable stilbenes are compounds such as resveratrol (trans-3,4′,5-trihydroxystilbene), abundant in grapes and red wine are known for their oxidative stress reducing, immune regulating, and cancer fighting effects. Pterostilbene (trans-3,5-dimethoxy-4′-hydroxystilbene), a more bioavailable counterpart of resveratrol found in blueberries and grapes, exhibits enhanced pharmacokinetics. Piceatannol (trans-3,5,3′,4′-tetrahydroxystilbene), present in red wine and passion fruit, functions as a tyrosine kinase inhibitor with anticancer potential. Other derivatives, such as piceid (resveratrol glucoside), astringin (piceatannol glucuronide), ε-viniferin, pallidol (resveratrol dimers), and gnetin C (a resveratrol dimer found in melinjo seeds), demonstrate anti-angiogenic and chemopreventive effects. These stilbenes control epigenetic pathways, specifically methylation of DNA, alteration of histone, and regulation of microRNA, contributing to their therapeutic efficacy, causing programmed cell death, lowering angiogenesis, and inflammatory responses. 28, 29 Methoxy-stilbenes exhibit anticancer activity by modulating key receptors and enzymes linked with the biosynthesis of estrogen in breast cancer cells. 30 Finings indicated that methoxy-stilbenes efficiently suppressed the action of two key enzymes involved in estrogen synthesis (aromatase and 17HSD17β). They are also involved in suppressing the expression of Erα, thereby weakening estrogen signalling and preventing cancer cell proliferation. Whole grape extracts may have more therapeutic benefits than isolated stilbenes. MBE, a rich source of resveratrol, piceatannol, and pterostilbene, showed greater potency than resveratrol by stimulating apoptosis and enforcing cell cycle arrest. 31 Resveratrol combats photo-carcinogenesis by adjusting apoptotic inhibition proteins and governing cell cycle signalling cascades. Stilbenes prevent prostate cancer through inhibition of MTA1 signaling pathways. Resveratrol and pterostilbene suppressed MTA1 expression, leading to reduced tumor growth, metastasis, and angiogenesis while enhancing apoptosis. 32
Carotenoids
Carotenoids are lipophilic pigments classified into carotenes and xanthophylls, which contribute to the red, yellow, and orange colours in fruits and vegetables. Beyond their roles as antioxidants, anti-inflammatories, and provitamin A sources, carotenoids demonstrate significant anticancer activity through mechanisms such as free radical neutralization, gene regulation, and modulation of inflammatory pathways. Key dietary sources of carotenoids include algae, yeast, carrots, tomatoes, citrus fruits, and various plant by-products. 33 Saponified lipophilic extract from sea buckthorn (Hippophae rhamnoides L.) has anticancer effects against breast cancer cell lines T47D and BT-549. 34 The extract inhibited proliferation (IC50 value: 16 µM), reduced ROS levels, and induced apoptosis (80.29% in T47D and 40.6% in BT-549). Carotenoids derived from yeast through cost-effective fermentation processes, demonstrating dose-dependent cytotoxicity against MCF7 and MDA-MB-231 cells (IC50 values: 29.11 and 7.82 μg/mL) while sparing normal HEK293T cells. 35 Similarly, carotenoids derived from Paracoccus sp. EGY7, identifying zeaxanthin (48.3%) as the predominant compound. The extract exhibited cytotoxicity against MDA-MB-231 cells (IC50 value: 1200 µg), inhibited cellular migration, and induced apoptosis through modulation of BAX/BCL2 ratios. 36 α-Carotene suppresses metastasis by modulating MMP2, MMP2, TIMP-1, and PAI-1, thereby impairing integrin β1/FAK/MAPK signaling without affecting primary tumor growth. β-Carotene inhibits gastric cancer progression through Notch/EMT pathway suppression, and in neuroblastoma, it downregulates MMPs, HIF-1α, and VEGF, consequently reducing metastasis. Additionally, it modulates M2 macrophage and fibroblast activation, limiting invasiveness in colorectal cancer. Overexpression of β-carotene 15,15′-oxygenase suppresses neuroblastoma self-renewal and metastasis through regulation of EMT and MMPs. 37
Major mechanistic anti-cancer potential of phytochemicals
Anti-angiogenesis and metastasis
Angiogenesis is the process by which old blood vessels are converted into new ones, crucial for tumor growth, invasion, and dissemination. Phenolic compounds serve as angiogenesis blockers to inhibit the multiplication of tumor cells. Polyphenol compounds such as EGCG, ellagic acid, and genistein counteract angiogenesis by inhibiting VEGF, PDGF, HIF-1α, and MMPs. Inhibition of platelet-derived growth factor (PDGF) receptor phosphorylation and epidermal growth factor receptor also aids in anti-angiogenesis. 38 Green tea polyphenols chelate ferrous ions and stop the proliferation of cancer cells driven by HIF 1α. In vitro studies on prostate tumor cells showed that EGCG treatment reduced HIF 1α-mediated transcription and protein levels under normal oxygen conditions. 39 Galanin, kaempferol, myricetin, and quercetin inhibit the tubular development of HUVECs mediated by vascular endothelial growth factor and impede U937 cell adherence to HUVECs. 40 Flavonoids, including kaempferol, genistein, apigenin, rutin, and naringin, reduce the levels of vascular endothelial growth factor in cells of human breast cancer. 14 Metastasis is the process by which cancer cells travel through lymph nodes to distant organs, interchangeable with angiogenesis, invasion, cell adhesion, migration, proteolysis, and extracellular matrix destruction. 41 Many dietary polyphenolic compounds disrupt tumor cell adhesion and migration, giving them anti-invasive and antimetastatic properties, though their precise signaling pathways and molecular mechanisms remain unidentified. 42 Curcumin inhibits VEGF/VEGFR2 signalling and downstream pathways (AKT, MAPK), reducing endothelial cell proliferation and migration. It also suppresses MMP2/9 expression and EMT by modulating Wnt/β-catenin pathways. 43 Resveratrol downregulates cyclin D1/CDK4 and VEGF expression, blocking endothelial cell activation, while Quercetin hinders CAM assay angiogenesis by targeting VEGF and HIF-1α. 44 Apigenin blocks STAT3 nuclear translocation, impairing endothelial tubulogenesis, while Genistein induces apoptosis in VEGF-loaded endothelial cells by inhibiting JNK/p38 and MMP-2/9. Luteolin downregulates PURB-mediated MAPK/PI3K pathways, reducing NSCLC angiogenesis. 45
Cell Cycle Arrest and Apoptosis
Apoptosis, a prevalent form of regulated cell death, is a critical target for various cancer therapies. Dietary cancer-fighting agents such as apigenin, chrysin, curcumin, EGCG, ellagic acid, resveratrol, and silymarin suppress carcinogenesis through the induction of apoptosis. 46 Notably, cancer cells exhibit heightened sensitivity to these agents compared to normal healthy cells. In sarcoma cells, EGCG induces apoptosis through mechanisms including cell cycle cessation at the G2/M phase, suppression of BCL-2, and the activation of p53 and BAX. 47 In prostate cancer cells, apoptosis is accompanied by the overexpression of BAX, along with the activation of caspases (3 and 8). EGCG primarily triggers and promotes senescence and apoptosis through p53-mediated signaling, in conjunction with the role of BAX. 48 Theaflavin, a phenolic compound in black tea, contributes to apoptosis by enhancing DNA fragmentation, activating caspases (3 and 8), increasing and decreasing the levels of BCL-2. 49 Furthermore, theaflavin activates the expression of caspases 9 and 3 in prostate cancer cells, alters the BAX/BCL-2 ratio, increases the release of cytochrome c from mitochondria, and causes apoptosis via p53 expression. 50 Anthocyanins, a significant class of flavonoids, demonstrated dose-related suppression of the growth of colon cancer cells. They induce programmed cell death in colon cancer cells through DNA damage and an imbalance in BAX and BCL-2 expression. However, certain bioflavonoids, including epicatechin, rutin, chlorogenic acid, and p-hydroxybenzoic acid, do not exhibit growth-inhibiting effects. 51 Delphinidin, a naturally occurring anthocyanidin, obstructs VEGF-induced cell movement and growth by arresting the cell cycle at the G0/G1 phase. The levels of Cyclin D1 and cyclin A significantly decreased. 52 Additionally, early activation of extracellular signal-regulated protein kinase 1/2, overexpression of caveolin-1, and downregulation of Ras contribute to the antiproliferative effects of delphinidin. 53
Modulation of cell signalling pathways
Phytochemicals exhibit various biological activities like anti-inflammatory, antiproliferative, antioxidant, antimutagenic, and immunomodulatory, which help regulate cancer progression and influence cancer development by modulating multiple signaling pathways. 54 They modulate routes concerned with cancer progression and suppression, such as the PI3K/Akt/mTOR/P70S6K pathway, PPARs, Nrf2, JAK-STAT, HIF-1, TGF-β, and TLR/NLRP, as well as the MAPK, ERK, and p38 pathways. 55-57 Phytochemicals can scavenge free radicals and respond to chemical stress, activating or inhibiting diverse signaling responses. Notable phytochemicals like apigenin, betulinic acid, ascorbic acid, curcumin, resveratrol, lycopene, and sesamol have protective effects through these pathways. 58 Resveratrol decreases the capacity of colorectal cancer cells to develop and metastasize. The mechanism indicates a connection between the modulation of the peroxisome proliferator gamma receptor coactivator 1-alpha signaling pathway and AMP-activated protein kinase. 59 Resveratrol suppresses ERK1/2 phosphorylation, limits gene expression associated with tumor progression. Resveratrol and curcumin act on Ras mutations that cause continuous activation of the MEK/ERK/MEK/Raf/Ras pathway, driving cell growth and survival. 60 Flavonoids such as luteolin, apigenin, and kaempferol inhibit AKT/mTOR/P13K signalling pathway, essential for cell survival, growth, and metastasis, while promoting apoptosis by suppressing AKT phosphorylation and modulating downstream regulators (STAT3). 61 Another dietary flavonoid, Fisetin, in combination with 5-FU treatment reduced PI3K expression, AKT activation, mTOR, its downstream targets, and mTOR signaling complex in PIK3CA-mutant cells. 62 Table 1 provides an overview of the regulatory pathways modulated by phytochemicals and their impact on cancer metabolism.
Table 1: Regulatory Pathways Modulated by Phytochemicals in Cancer Metabolism
| Pathway | Phytochemicals | Mechanism of action | Effect on Cancer Metabolism |
| PI3K/AKT/mTOR
One of the most frequently dysregulated signaling cascades in cancer, promoting cell survival, proliferation, and resistance to apoptosis |
Curcumin, Resveratrol, Luteolin, Apigenin | Inhibits Akt phosphorylation; suppresses mTOR and downstream effectors (FOXO, STAT3) | Reduces cell survival, proliferation, and metastasis; enhances chemo-sensitivity. 40 |
|
MAPK/ERK (Ras/Raf/MEK/ERK) Regulates the growth of cells. Its dysfunction is linked to uncontrolled tumour growth. |
Resveratrol, Ursolic Acid, Gingerol | Blocks ERK1/2 phosphorylation; inhibits Ras/Raf activation | Suppresses uncontrolled proliferation and metastasis; induces apoptosis. 63,64 |
| NF-κB
Key regulator of inflammation-driven tumorigenesis |
Curcumin, Quercetin, Ellagic Acid | Inhibits NF-κB nuclear translocation and DNA binding | Reduces inflammation-driven tumorigenesis and angiogenesis. 61 |
| Wnt/β-catenin | Curcumin, Genistein | Downregulates Wnt signaling; suppresses β-catenin nuclear accumulation | Inhibits EMT and metastasis. 65, 66 |
| Apoptosis (Intrinsic) | Luteolin, Icanin | Modulates BCL-2/BAX ratio; activates caspases via mitochondrial ROS | Triggers caspase-dependent apoptosis; reduces cancer cell survival. 67, 68 |
| Apoptosis (Extrinsic) | Irigenin, Apigenin, Kaempferol | Activates TRAIL/DR5 and FADD pathways | Enhances death receptor-mediated apoptosis. 69 |
| ROS/Nrf2-ARE | Phenolic compounds | Activates Nrf2-ARE pathway; upregulates SOD, CAT, GPX | Counteracts oxidative stress; promotes detoxification and chemoprevention. 70, 71 |
|
p53 Suppressor pathway regulates cell cycle and apoptosis |
Ellagic Acid, Resveratrol | Upregulates p53 and p21; downregulates cyclins (D1, E) | Induces G0/G1 cell cycle arrest; restores tumor-suppressive functions. 72, 73 |
| Hormone Receptors | Genistein, Apigenin | Modulates ER/AR signaling; inhibits HER2/STAT3 | Suppresses hormone-driven proliferation. 74, 75 |
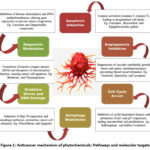 |
Figure 1: Anticancer mechanism of phytochemicals: Pathways and molecular targets |
In 2020, GLOBOCAN reported 19.3 million new cases of cancer worldwide, ranking India third behind the US and China. Projections indicate that by 2040, the percentage of people suffering from cancer in India will rise to 2.08 million, reflecting a 57.5% increase from 2020. 76, 77 Cancer can develop in nearly any organ or system within the human body, including the oral cavity, breast, bone, brain, stomach, intestines, lungs, renal organs, genitals, and the reproductive system. According to a recent review, the five most common cancer locations across various anatomical sites in Indian men include the stomach (4.8%), prostate (6.1%), tongue (5.9%), mouth (8.4%), and lung (10.6%). For women, the most commonly reported cancer sites were breast (28.8%), cervix (10.6%), ovary (6.2%), corpus uteri (3.7%), and lung (3.7%). Furthermore, based on estimated incidence rates, cumulative risk, and total cancer burden across all anatomical sites, the most prevalent cancers were those of the digestive system (1 in 39 individuals), breast (1 in 56), genital system (1 in 53), oral cavity (1 in 62), and respiratory system (1 in 74). Among digestive system cancers, the most frequently diagnosed malignancies included the stomach (1 in 213), liver (1 in 276), and colorectal cancer (1 in 295). In contrast, lung cancer remained the predominant malignancy in the respiratory system (1 in 101). 78 The anticancer potential of phytochemicals derived from various food sources is supported by animal and human studies. Tables 2-6 present their correlation with a reduced incidence of conditions such as breast, lung, colon, stomach, liver, and prostate cancer, along with the underlying mechanisms of their anticancer activity. These tables also highlight the effectiveness of phytochemicals in cancer prevention and mitigation.
Table 2: Anticancer effects of phytochemicals in various animal models
| Phytochemical | Type of cancer | Action |
| Apigenin – flavonoids | NSCLC | Regulates Bcl-2 family protein levels and triggers caspase activation, resulting in G2/M phase cell cycle arrest and apoptotic cell death in NSCLC xenograft models. 79 |
|
Epigallocatechin – flavonoids
|
Breast cancer | Hinders tumor development and decreases tumor burden by triggering apoptosis and restraining the growth of human breast cancer cells in a mouse model, in addition to mitigating nitrosamine-induced lung tumor formation. 80 |
| Prostate cancer | Regulates essential signaling pathways in multiple tumor models, including MAPK and PI3K/AKT, demonstrating anti-angiogenic, anti-proliferative, and pro-apoptotic effects. 81 | |
| Gingerol – Phenolic compound | Breast cancer | Limits the expansion and metastatic potential of 4T1Br4 mammary carcinoma in a syngeneic breast cancer model. Reduces proliferation and invasiveness of lung-metastatic breast cancer cells by inhibiting Akt, p38MAPK, and EGFR pathways. 82 |
| Curcumin – Phenolic compound | Colon cancer | Reduces the expression of the K-RAS and β-catenin genes, COX-2, and survivin within colon tissue in Sprague-Dawley rat models. 83 |
| Resveratrol and Pterostilbene – Stilbenes | Prostate cancer | Inhibits MTA1-regulated miR-17 family, miR-22, and miR-34a, resulting in reduced prostatic hyperplasia, tumor growth, and the proliferation and invasion of prostate cancer cells in murine models. 32 |
|
β-sitosterol – Phytosterol
|
Colon cancer | Mitigates the decline of antioxidant enzymes and reestablishes nonenzymatic antioxidant defense in a rat model of colon cancer induced by 1,2 DMH. 84 |
| Lutein – Carotenoid |
Colon cancer |
Suppresses K-RAS and AKT expression and reduces aberrant crypt foci in a DMH-induced mouse model of colon cancer. 85 |
Table 3: Cancer protective properties of whole grains
| Functional compounds | Cancer
type |
Mechanisms | Clinical evidence | |
| Study design | Findings | |||
|
Alkylresorcinols, Avenanthramide, anthocyanins, lignans, flavones, ferulic acid, gallic acid, dietary fiber, |
Stomach |
Maintains gut microbiota and inhibits the action of Helicobacter pylori Suppression of cell metastasis, cell growth, and promotion of Induction of apoptosis |
Meta-analysis of CCS using the highest vs the lowest intake comparison | Reduced whole grain intake coupled with high intake of refined grains contributed to a 36% higher risk of gastric cancer. 86 |
| Meta-analysis of 1 longitudinal study and 2 CCS | Daily intake of whole grains decreased the risk of gastric cancer by 65%. 87 | |||
| Liver | Reduces inflammation and oxidative stress
Mitigates the carcinogenic effect of hyperinsulinemia |
Dose-dependent meta-analysis of 6 CS | A daily intake of 50g of whole grains decreased liver cancer risk by 23%. 88 | |
| Colorectal | Increases the bulk of fecal matter
Decreases the transit time and prevents carcinogenic compounds from being absorbed Butyrate, a short-chain fatty acid, promotes apoptosis |
Dose-dependent meta-analysis of 111 CS | Daily intake of 90g of whole grains decreased colorectal cancer risk by 17%. 89, 90 | |
| Lung | Reduces inflammation | A prospective cohort study with 101,732 participants | Higher whole grain intake was associated with a 16% reduced risk of lung cancer. 91 | |
| Breast | Upregulation of apoptosis
Inhibiting the action of the MMP–9 and MMP-2 Reduces the action of urokinase-type plasminogen activator Down-regulation of STAT5/IGF-1R and STAT3/VEGF pathways |
Meta-analysis of CS and 7 CCS | A daily intake of 50g of whole grains decreased breast cancer risk by 17%. 92 | |
| Prostate | Reduces the levels of inflammatory biomarkers (IL 1β, TNF α, and 5-lipoxygenase)
Increases the apoptotic factors (pro-apoptotic caspase 3 protein) |
CCS (120 participants) | Greater adherence to a healthy diet and a higher phytochemical index were associated with reduced odds of prostate cancer (0.322). 93 | |
Table 4: Association between red and processed meat consumption and cancer risk
|
Cancer type |
Mechanisms | Clinical evidence | |
| Study design | Findings | ||
| Stomach
Liver and Breast cancer |
H. pylori infection causes stomach cancer via two pathways
Direct pathway: Induction of protein modulation and mutation of specific genes Indirect pathways: Inflammation of gastric epithelial cells. Interference of CagA with certain host proteins involved in the cell cycle and motility Heme iron in red meat causes the production of cancer-causing N-nitroso compounds, which lead to DNA damage and cellular oxidative stress. |
Meta-analysis of 232 studies (33,831,063 participants) |
H. pylori colonization increased stomach cancer risk by 2.56 times (95% CI, 2.18 – 3.00). 94 Red meat intake increased stomach cancer risk in H. pylori individuals (OR=1.85). 95 |
| Systematic review and meta-analysis of 148 prospective studies | Intake of red meat may increase the risk of breast cancer. Processed meat increases the risk of liver and breast cancer by 6% and 12%. 96, 97 | ||
| Colorectal | KRAS, TP53, and APC mutations
Heme iron in red meat promotes the formation of cancer-causing compounds (HCAs and PAHs) |
Systematic review and meta-analysis of 40 CS | High intake of red (RR: 1.09) and processed meat (RR: 1.19) raises the risk of colorectal cancer. Daily intake of 76g of red and processed meat increases the risk of colorectal cancer by 20%. 98, 89 |
| Prostate | Method of cooking – grilling and barbecuing, and release of nitrates, nitrites, and other carcinogenic compounds | Meta-analyses of 72 studies | Red meat intake was not linked with prostate cancer. However, consumption of processed meat increased the risk of prostate cancer. 100 |
Table 5: Cancer protective mechanisms of fruits and vegetables
| Functional compounds | Cancer
type |
Mechanisms | Clinical evidence | |
| Study design | Findings | |||
|
Lycopene, flavonols, Lignan Quercetin, Resveratrol Geinstein Flavonols Kaempferol Anthocyanins, Proanthocyanidins, Punicalagin, Glucosinolate, isothiocyanates, carotenoids, dietary fibre |
Breast
Liver |
Lycopene demonstrated antiproliferative activity by inhibiting the multiplication of insulin-like growth factor-I-stimulated cells Dietary fibre in fruits/dried fruits interacts with oestrogen metabolism, thereby reducing its bioavailability Lignan binds with oestrogen receptors, reducing endogenous oestrogen levels Induce apoptosis Stops the G2/M phase of the cell cycle Reduces the levels of EMT-related markers and disrupts the PI3K/AKT pathway |
A prospective CS | High intake of tomatoes decreased the risk of breast cancer (RR: 0.87), and high intake of dried fruits decreased endometrial cancer risk (RR: 0.60).101 |
| A prospective CS | High consumption of fruits and vegetables (>5.5 servings/day), especially cruciferous, yellow-, and orange-coloured vegetables, decreased the risk of breast cancer. 102 | |||
| Meta-analysis of 9 CS(N=13,26,176)
|
Participants who ate more vegetables had a 39% lower chance of developing liver cancer. 103, 104 | |||
| Lung | Inhibits cancer cell growth and proliferation
Promotes cell apoptosis, Reduces cell migration and adhesion, and sensitizes cancer cells to antitumor drugs |
Meta-analysis of 18 prospective studies | Participants who consumed more fruits and vegetables had a lower risk of lung cancer than their counterparts. (RR: 0.92 for vegetables, and 0.82 for fruits).105 | |
| Colorectal | Reduces the levels of CFL2, which is responsible for promoting actin filament and reducing cell stiffness, thereby reducing colon cancer risk
Increases the release of several proinflammatory cytokines from immune cells, thereby promoting anticancer cytotoxicity |
Meta-analyses of 24 observational studies (10,68,158 participants) | Increased consumption of citrus fruits, apples, watermelon, and kiwi decreased colorectal cancer risk by 9%, 25%, 26%, and 13%, with corresponding RR of 0.91, 0.75, 0.74, and 0.87, respectively.106-108 | |
Table 6: Cancer protective potential of nuts, oilseeds, and tea
| Functional compounds | Cancer
type |
Mechanisms | Clinical evidence | |
| Study design | Findings | |||
| Nuts and oilseeds | ||||
|
Hydroxybenzoic acid, flavones, flavanols, hydroxycinnamic acid; flavonols, catechins, flavanones, stilbenes, anthocyanins, isoflavones, lignans, ellagic acid, Phytosterols, SITO phytostanols, STIG, phytostanol, ATA, ERGO, CAMP. |
Prostate | Alters the levels of inflammatory markers, including C-reactive protein, ILs, TNF-α, and cell adhesion molecules
Contributes to antioxidant activity |
Systematic review and dose-dependent meta-analysis | Consumption of nuts was not associated with prostate cancer risk. 109-110 |
| Colon | Reduce DNA damage via down-regulation of inflammation gene expression (Hong et al., 2021)
Decrease PG E2 production, IL-6 and IL-8 release, and COX – 2 expression |
Meta-analyses of 8 CS and 5 CCS | Daily nut consumption of 28g may offer a protective effect, lowering colorectal cancer risk by 33%. 111 | |
| Tea | ||||
| Polyphenols, EGCG, epicatechin, epigallocatechin, gallocatechin gallate, epigallocatechin | Colorectal | Polyphenols modulate multiple signaling pathways involved in cancer cell proliferation and apoptosis, including the MAPK, PI3K/Akt, Wnt/β-catenin, and 67 kDa laminin receptor pathways
Inhibit new blood vessel formation (angiogenesis) |
Meta-analysis on 14 studies | Tea was associated with a 24% reduction in colon cancer risk; however, considerable heterogeneity was observed, with estimates ranging from a 51% decrease to an 18% increase, suggesting the influence of population and regional differences. 112 |
Challenges in the bioavailability of phytochemicals
Phytochemicals provide a wide range of health benefits, but their full therapeutic potential is often affected by factors such as limited bioavailability and stability. After oral ingestion, several processes, including digestion, absorption, metabolism, distribution, and excretion, affect the bioavailability of phytochemicals.113 The structural and compositional characteristics of food matrices are crucial in determining how phytochemicals are released during digestion. These compounds are entrapped within the cell walls and influence their bioaccessibility. Food processing techniques, particularly thermal treatments, cause the release of these functional compounds by disrupting their natural cell structure. 114 These processing techniques also result in degradation of thermolabile compounds due to oxidative and hydrolytic reactions. The natural physical and chemical properties, such as crystallinity and fat solubility, determine the absorption and bioavailability. The chemical traits, such as hydrophobicity, molecular weight, can impede their solubility and transport across the gastrointestinal epithelium. Physiological factors such as changes in gastrointestinal pH, enzymatic functions, and the presence of specific transport systems also impact their absorption rate. Antinutritional factors, including phytates, tannins, and oxalates, interact with phytochemicals and form complexes, thus diminishing their bioavailability. Dietary fibers hinder the absorption of fat-soluble compounds by modifying their interaction with lipid micelles. 115 After the process of absorption, metabolism either enhances or decreases their bioactivity of phytochemicals. This metabolic process is influenced by gut microbiota, which can affect their chemical structures and biological functions. 116
Table 7 presents an overview of phytochemical bioavailability, highlighting its challenges, underlying mechanisms, and strategies for enhancement.
Table 7: Phytochemical Bioavailability: Challenges, Mechanisms, and Enhancement Strategies
| Phytochemicals | Challenges faced in bioavailability | Strategies to enhance the bioavailability of phytochemicals | |
| Flavonoids | Quercetin, Kaempferol, Naringenin, Catechins | Glycosylation slows absorption, competition for intestinal transporters, and rapid excretion. 117 | Enzyme-based hydrolysis (to convert glycosides to aglycones), co-administration with inhibitors of efflux transporters, and nanoformulations. 118 |
| Phytosterols | β-Sitosterol, Campesterol, Stigmasterol | Phytosterols are highly hydrophobic, limiting their solubility in aqueous environments, leading to poor gastrointestinal absorption. Phytosterols are prone to oxidation, and their effect is reduced. 119 | Use of lipid-based delivery systems such as emulsions, micelles, or nanoencapsulation to improve solubility and enhance bioavailability. Use of microencapsulation techniques, like spray drying, to protect against oxidation. 120 |
| Polyphenols | Curcumin, Resveratrol, Quercetin, EGCG, and Anthocyanins | Poor water solubility, chemical instability in gastric pH, rapid metabolism (glucuronidation and sulfation), and low intestinal absorption. 121 | Nano emulsions, liposomal encapsulation, polymeric nanoparticles, and co-administration with bioavailability enhancers. 122 |
| Stilbenes | Resveratrol, Pterostilbene | Low aqueous solubility, leading to poor gastrointestinal absorption and bioavailability. Rapid oxidation and photodegradation reduce the stability of stilbenes, limiting their effectiveness.123 | Use lipid-based formulations such as nano emulsions, liposomes, or solid lipid nanoparticles to improve solubility and bioavailability. Encapsulation techniques can be used to prevent oxidation. 124 |
| Carotenoids | β-Carotene, Lycopene, Lutein | Poor water solubility, degradation in heat and light, and reduced absorption due to binding with fiber. 125 | Lipid-based delivery systems, microencapsulation, and dietary co-administration with fats. 126 |
Discussion
Cancer remains one of the most challenging NCDs to treat due to its complex pathophysiology, heterogeneity, and ability to develop resistance to conventional therapies. Chemotherapy is associated with severe side effects, including systemic toxicity and damage to non-cancerous cells. In this context, incorporating naturally derived bioactive compounds into cancer treatment regimens has emerged as a highly promising strategy. The integration of phytochemicals with conventional chemotherapeutic agents has garnered significant attention in recent years.
This review provides an in-depth over view of the role of specific phytochemicals, particularly flavonoids, phenolic acids, phytosterols, stilbenes, and carotenoids in preventing and treating cancer. These non-nutritive compounds found in whole grains, fruits, vegetables, and leaf extracts exhibit antioxidant, anti-inflammatory, and anticancer effects. Their anticarcinogenic effects are mainly mediated through the modulation of critical intracellular signaling pathways involved in cancer pathophysiology. Key signaling cascades influenced by these non-nutritive compounds include the PI3K/Akt/mTOR pathway, which controls cell growth, survival, and metabolism; the NF-κB signaling pathway, central to inflammation and immune response; and the MAPK/ERK pathway, which controls cell proliferation and differentiation. Furthermore, they play an essential role in regulating and maintaining the autophagy-apoptosis balance. Phytochemicals exert a multi-targeted mode of action by interacting with various molecular targets simultaneously, in contrast to conventional chemotherapeutic agents, which exhibit single pathway specificity while destroying cancer cells. The potential of phytochemicals to simultaneously disrupt several signalling cascades improves cancer treatment.
Extensive preclinical studies using in vitro and in vivo models have demonstrated the anticancer effects of these phytochemicals across a range of cancer types, including breast, colon, prostate, lung, and liver cancers. Moreover, clinical trials and meta-analyses of cohort and prospective studies have provided epidemiological evidence supporting the protective role of diets rich in different phytochemicals. These findings highlight the potential of plant-based diets as adjuncts or alternatives to conventional cancer therapies, emphasizing the significance of dietary interventions in cancer prevention and therapy.
Conclusion
Cancer continues to pose a significant global health burden, necessitating the development of cost-effective, safe, and widely accessible therapeutic strategies. Increasing attention is being directed toward natural products due to their easy accessibility and affordability. Among these natural compounds, phytochemicals have emerged as promising compounds for anticancer drug development. This review offers a comprehensive examination of key phytochemicals, particularly flavonoids, phenolic acids, stilbenes, phytosterols, and carotenoids as anticancer agents. The underlying mechanisms of action, including modulation of critical signaling pathways, apoptosis induction, anti-inflammatory effects, and antioxidative activity, are discussed in detail, supported by evidence from both preclinical studies and human clinical trials.
Clinical trials remain the gold standard for evaluating therapeutic efficacy and safety. While meta-analyses of cohort and prospective studies have highlighted the potential anticancer effects of dietary phytochemicals against various cancer types, several limitations persist. These include variability in the phytochemical content of foods, influenced by factors such as plant variety, cultivation conditions, processing methods, and challenges related to bioavailability. The latter is particularly critical, as the absorption, metabolism, and systemic availability of phytochemicals can significantly impact their biological efficacy.
Although numerous experimental studies have explored strategies to enhance phytochemical bioavailability, such as nano formulations, co-administration with bioenhancers, and structural modifications, clinical research on the pharmacokinetics, potential nutrient interactions, optimal dosing, and long-term safety of isolated or enriched phytochemicals remains limited. Further well-designed human studies are essential to translate promising laboratory findings into effective, evidence-based interventions for cancer prevention and therapy.
Acknowledgement
The authors acknowledge the institutional support (M.O.P. Vaishnav College for Women, Autonomous), Chennai, for providing access to scientific databases and academic resources that helped in the compilation of information.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article
Conflict of Interest
The authors do not have any conflicts of interest.
Data Availability Statement
Although adequate data have been presented in tables and figures, all authors affirm that any additional data will be provided upon request if needed.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to Reproduce Material from Other Sources
Not applicable
Clinical Trial Registration
This research does not involve any clinical trials.
Author Contributions
- Sarah Jane Monica: Conceived the idea, drafted the manuscript, supervised, reviewed, edited, and enhanced the overall quality of the manuscript
- Deevena Jemima: Conceived the idea, prepared tables, and contributed to drafting the manuscript.
- Esther Lydia D: Created figures, supervised, and contributed to drafting the manuscript.
- Priyadarshini S: Prepared tables and contributed to drafting the manuscript.
- Anne Mary Preetha: Prepared tables and contributed to drafting the manuscript.
- Emi Grace Mary Gowshika Rajendran: Contributed to drafting the manuscript.
References
- Bhuiyan M. A., Galdes N., Cuschieri S.H.P. A comparative systematic review of risk factors, prevalence, and challenges contributing to non-communicable diseases in South Asia, Africa, and Caribbeans. J Health Popul Nutr. 2024; 43: 140. DOI: https://doi/org/10.1186/s41043-024-00607-2.
CrossRef - Sucharitha A., Bhuvana D. An overview of cancer. Adv Cancer Chemother Pharmacol. 2023; 1(1): 16000102. DOI: https://doi/org/10.23880/accp-16000102.
CrossRef - Sung H., Ferlay J., Siegel R.L., Laversanne M., Soerjomataram I., Jemal A., Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: Cancer J Clin. 2021; 71(3): 209-249. DOI: https://doi/org/10.3322/caac.21660.
CrossRef - Debela D.T., Muzazu S.G., Heraro K.D., Ndalama M.T., Mesele B.W., Haile D.C., Kitui S.K., Manyazewal T. New approaches and procedures for cancer treatment: Current perspectives. Sage Open Med. 2021; 12:9 DOI: https://doi/org/10.2050312121103436.
CrossRef - Rudzinska A., Juchaniuk P., Oberda J., Wisniewska J., Wojdan W., Szklener K. Phytochemicals in cancer treatment and cancer prevention-review on epidemiological data and clinical trials. Nutr. 2023; 15(8): 1896. DOI: https://doi/org/10.3390/nu15081896.
CrossRef - Paul J. K., Azmal M., Haque S.N.B., Talukder O.F., Meem M., Ghosh A. Phytochemical-mediated modulation of signalling pathways: A promising avenue for drug discovery. Adv Redox Res. 2024; 13: 100113. DOI: https://doi.org/10.1016/j.arres.2024.100113
CrossRef - Rathee S., Patil U.K., Jain S.K., Exploring the potential of dietary phytochemicals in cancer prevention. A comprehensive review. J Explor Res Pharmacol. 2024; 9(1): 34-47. DOI: https://doi/org/10.14218/JERP.2023.00050.
CrossRef - George B.P., Chandran R., Abrahamse H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants (Basel). 2021; 14; 10(9): 1455. DOI: https://doi/org/10.3390/antiox10091455.
CrossRef - Bahrami H. Nutrition for cancer prevention and control. A review of dietary risk factors and protective measures. Cancer Screen Prev. 2023; 2(4): 250-259. DOI: https://doi/org/10.14218/CSP.2023.00008S.
CrossRef - Bahrami H., Tafrihi M., Mohamadzadeh S. Reversing the Warburg effect to control cancer: A review of diet-based solutions. J Curr Oncol Med Sci. 2022; 2(3):234-248.
- Hardt L., Mahamat-Saleh Y., Aune D., Schlesinger S. Plant-based diets and cancer prognosis: A review of recent research. Curr Nutr Rep. 2022; 11(4):695-716. DOI: https://doi/org/10.1007/s13668-022-00440-1
CrossRef - Choudhari A.S., Mandave P.C., Deshpande M., Ranjekar P., Prakash O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front Pharmacol. 2020; 10: 1614. DOI: https://doi.org/10.3389/fphar.2019.01614
CrossRef - Cai J., Tan X., Hu Q., Pan H., Zhao M., Guo C., Zeng J., Ma X., Zhao Y. Flavonoids and Gastric Cancer Therapy: From Signaling Pathway to Therapeutic Significance. Drug Des Devel Ther. 2024; 25; 18: 3233-3253. DOI: https://doi/org/10.2147/DDDT.S466470
CrossRef - Slika H., Mansour H., Wehbe N., Nasser S.A., Iratni R., Nasrallah G. Shaito A., Ghaddar T., Kobeissy F., Eid AH. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed Pharmacother. 2022; 146: 112442. DOI: https://doi.org/10.1016/j.biopha.2021.112442
CrossRef - Saleh A., ElFayoumi H, M., Youns M., Barakat W. Rutin and orlistat produce antitumor effects via antioxidant and apoptotic actions. Naunyn Schmiedebergs Arch Pharmacol. 2019; 392: 165-175. DOI: https://doi/org/10.1007/s00210-018-1579-0
CrossRef - Hu X., Li J., Fu M., Zhao X., Wang W. The JAK/STAT signaling pathway: from bench to clinic. Sig Transduct Target Ther. 2021; 6(1): 402. DOI: https://doi/org/10.1038/s41392-021-00791-1
CrossRef - Zhang Y., Lin H., Liu C., Huang J., Liu Z., A review for physiological activities of EGCG and the role in improving fertility in humans/mammals. Biomed Pharmacother. 2020; 27: 110186. DOI: https://doi.org/10.1016/j.biopha.2020.110186
CrossRef - Naponelli V., Rocchetti M, T., Mangieri D. Apigenin: Molecular mechanisms and therapeutic potential against cancer spreading. Int J Mol Sci. 2024; 25(10): 5569. DOI: https://doi.org/10.3390/ijms25105569
CrossRef - Jiang L., Zhao X., Xu J., Li C., Yu Y., Wang W., Zhu L. The protective effect of dietary phytosterols on cancer risk: A Systematic Meta-Analysis. J Oncol. 2019; 23: 7479518. DOI: https://doi/org/10.1155/2019/7479518
CrossRef - Zhao J. Q., Hao Y. Y., Gong T. T., Wei Y. F., Zheng G., Du Z. D., Zou B.J., Yan S., Liu F.H., Goa S., Wu, Q.J., Zhao YH. Phytosterol intake and overall survival in newly diagnosed ovarian cancer patients: An ambispective cohort study. Front Nutr. 2022; 25 (9): 974367. DOI: https://doi/org10.3389/fnut.2022.974367
CrossRef - Banerjee J., Karati D. In-silico pharmacology against cancer of a novel phytosterol dendrosterone isolated from plant Dendrobium ochreatum. Pharmacol Res Mod Chin Med. 2022; 4: 100149. DOI: https://doi/org/10.1016/j.prmcm.2022.100149
CrossRef - Gu S., Liu F., Xie X., Ding M., Wang Z., Xing X. Xiao T., Sun X. β-Sitosterol blocks the LEF-1-mediated Wnt/β-catenin pathway to inhibit proliferation of human colon cancer cells. Cell Signal. 2023; 104: 110585. DOI: https://doi.org/10.1016/j.cellsig.2022.110585.
CrossRef - Liu J., Han X., Zhang T., Tian K., Li Z., Luo F. Reactive oxygen species (ROS) scavenging biomaterials for anti-inflammatory diseases: from mechanism to therapy. J Hematol Oncol. 2023; 16(1):116. DOI: https://doi/org/10.1186/s13045-023-01512-7
CrossRef - Jomova K., Raptova R., Alomar S.Y., Alwasel S.H., Nepovimova E., Kuca K., Valko M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol. 2023; 97(10): 2499-2574. DOI: https://doi/org/10.1007/s00204-023-03562-9
CrossRef - Abotaleb M., Liskova A., Kubatka P., Busselberg, D. Therapeutic potential of plant phenolic acids in the treatment of cancer. Biomolecules. 2020; 10(2): 221. DOI: https://doi.org/10.3390/biom10020221
CrossRef - Assumpcao J.H.M., Takeda A.A.S., Sforcin J.M., Rainho C. A. Effects of propolis and phenolic acids on triple-negative breast cancer cell lines: potential involvement of epigenetic mechanisms. Molecules. 2020; 25(6): 1289.DOI: https://doi.org/10.3390/molecules25061289
CrossRef - Murai, T., Matsuda, S. The chemopreventive effects of chlorogenic acids, phenolic compounds in coffee, against inflammation, cancer, and neurological diseases. Molecules. 2023; 28(5): 2381. DOI: https://doi.org/10.3390/molecules28052381
CrossRef - Wang L.Y, Zhao S., Lv G.J, Ma X.J., Zhang J.B. Mechanisms of resveratrol in the prevention and treatment of gastrointestinal cancer. World J Clin Cases. 2020; 8(12): 2425-2437. DOI: https://doi.org/10.12998/wjcc.v8.i12.2425
CrossRef - Levenson AS. Dietary stilbenes as modulators of specific miRNAs in prostate cancer. Front Pharmacol. 2022; 24;13: 970280. DOI: https://doi/org/10.3389/fphar.2022.970280
CrossRef - Licznerska B., Szaefer H., Wierzchowski M., Mikstacka R., Papierska K., Baer-Dubowska W. Evaluation of the effect of the new methoxy-stilbenes on expression of receptors and enzymes involved in estrogen synthesis in cancer breast cells. Mol Cell Biochem. 2018; 44(1-2):53-62. DOI: https://doi/org/10.1007/s11010-017-3230-7
CrossRef - Balasubramani S.P., Rahman M.A., Basha S.M. Synergistic action of stilbenes in muscadine grape berry extract shows better cytotoxic potential against cancer cells than resveratrol alone. Biomed. 2019; 5;7(4):96. DOI: https://doi/org/10.3390/biomedicines7040096
CrossRef - Levenson A. S. Metastasis-associated protein 1-mediated antitumor and anticancer activity of dietary stilbenes for prostate cancer chemoprevention and therapy. Semin Cancer Biol. 2022; 80: 107-117. DOI: https://doi/org/10.1016/j.semcancer.2020.02.012
CrossRef - Bas T.G. Bioactivity and bioavailability of carotenoids applied in human health: Technological advances and innovation. Int J Mol Sci. 2024; 25(14):7603. DOI: https://doi/org/10.3390/ijms25147603
CrossRef - Visan S., Soritau O., Tatomir C., Baldasici O., Balacescu L., Balacescu O., Muntean P., Gherasim C., Pintea A. The bioactive properties of carotenoids from lipophilic sea buckthorn extract (Hippophae rhamnoides) in breast cancer cell lines. Molecules. 2023; 28; 11: 4486. DOI: https://doi.org/10.3390/molecules28114486
CrossRef - Sinha S., Das S., Saha B., Paul D., Basu B. Anti-microbial, anti-oxidant, and anti-breast cancer properties unraveled in yeast carotenoids produced via cost-effective fermentation technique utilizing waste hydrolysate. Front Microbiol. 2023; 13: 1088477. DOI: https://doi.org/10.3389/fmicb.2022.1088477
CrossRef - Abdelazim K., Hussein A., Khattab S. N., El Feky S. E., Noby, N. Impact of Paracoccus EGY7 carotenoids on triple-negative breast cancer cells: in vitro study. AMB Express. 2025; 15(1): 21. DOI: https://doi.org/10.1186/s13568-025-01825-5
CrossRef - Koklesova L., Liskova A., Samec M., Zhai K., Abotaleb M., Ashrafizadeh M., Brockmueller A., Shakibaei M., Biringer K., Bugos O., Najafi M., Golubnitschaja O., Busselberg D., Kubatka P. Carotenoids in cancer metastasis – Status quo and outlook. Biomolecules. 2020; 10(12); 1653. DOI: https://doi.org/10.3390/biom10121653
CrossRef - Cheng K., Liu C. F., Rao G., W. Anti-angiogenic Agents: A review on vascular endothelial growth factor receptor-2 (VEGFR-2) inhibitors. Curr Med Chem. 2021; 28(13): 2540–2564. DOI: https://doi.org/10.2174/0929867327666200514082425
CrossRef - Ma Z., Xiang X., Li S., Xie P., Gong Q., Goh B.C., Wang L. Targeting hypoxia-inducible factor-1, for cancer treatment: Recent advances in developing small-molecule inhibitors from natural compounds. Semin Cancer Biol 2022; 80: 379-390. DOI: https://doi.org/10.1016/j.semcancer.2020.09.011
CrossRef - Ahmed M.B., Islam S.U., Alghamdi A.A.A., Kamran M., Ahsan H., Lee Y.S. Phytochemicals as chemo-preventive agents and signalling molecule modulators: current role in cancer therapeutics and inflammation. Int J Mol Sci. 2022; 23(24): 15765. DOI: https://doi.org/10.3390/ijms232415765
CrossRef - Libby J.R., Royce H., Walker S.R., Li L. The role of extracellular matrix in angiogenesis: Beyond adhesion and structure. Biomat Biosyst. 2024; 15: 100097. DOI: https://doi.org/10.1016/j.bbiosy.2024.100097
CrossRef - Shah N.H., Kuriyan, J. Understanding molecular mechanisms in cell signaling through natural and artificial sequence variation. Nat Sci Mol Biol. 2019; 26, 25–34. DOI: https://doi.org/10.1038/s41594-018-0175-9
CrossRef - Khater M., Greco F., Osborn H.M.I. Antiangiogenic activity of flavonoids: A systematic review and meta-analysis. Molecules. 2020; 25(20): 4712. DOI: https://doi.org/10.3390/MOLECULES25204712
CrossRef - Liskova A., Koklesova L., Samec M., Smejkal K., Samuel S. M., Varghese E., Abotaleb M., Biringer K., Kudela E., Danko J., Shakibaei M., Kwon T.K., Busselberg D., Kubatka P. Flavonoids in cancer metastasis. Cancers. 2020; 12(6), 1498. DOI: https://doi.org/10.3390/CANCERS12061498
CrossRef - Lekhak N., Bhattarai H. K. Phytochemicals in cancer chemoprevention: Preclinical and clinical studies. Cancer Control. 2024; 31:10732748241302902. DOI: https://doi/org/10.1177/10732748241302902
CrossRef - Li C., Xu Y., Zhang J., Zhang Y., He W., Ju J., Wu Y., Wang Y. The effect of resveratrol, curcumin, and quercetin combination on immuno-suppression of tumor microenvironment for breast tumor-bearing mice. Sci Rep. 2023; 13278 DOI: https://doi.org/10.1038/s41598-023-39279-z
CrossRef - Belyaeva E., Loginova N., Schroeder B. A., Goldlust I.S., Acharya A., Kumar S., Timashev P., Ulasov I. The spectrum of cell death in sarcoma. Biomed Pharmacother. 2023; 162 (114683). DOI: https://doi.org/10.1016/j.biopha.2023.114683.
CrossRef - Wang H., Guo M., Wei H., Chen Y. Targeting p53 pathways: mechanisms, structures and advances in therapy. Sig Transduct Target Ther. 2023; 8: 92. DOI: https://doi.org/10.1038/s41392-023-01347-1
CrossRef - Dong S., Wu S., Hao F., Wu J., Liao Z., Zhong Q., Zhong R., Fang X. Research advancements on theaflavins: Isolation, purification, synthesis, gut microbiota interactions, and applications potentials. Food Res Int. 2025; 202 (115692). DOI: https://doi.org/10.1016/j.foodres.2025.115692
CrossRef - Czabotar P.E., Garcia-Saez A.J. Mechanisms of BCL-2 family proteins in mitochondrial apoptosis. Nature Reviews Mol Cell Biol. 2023; 24: 732–748. DOI: https://doi.org/10.1038/s41580-023-00629-4
CrossRef - Nwozo O.S., Effiong E.M., Aja P.M., Awuchi C. G. Antioxidant, phytochemical, and therapeutic properties of medicinal plants: A review. Int J Food Prop. 2023; 26(1), 359–388. DOI: https://doi.org/10.1080/10942912.2022.2157425
CrossRef - Sharma A, Choi HK, Kim YK, Lee HJ. Delphinidin and its glycosides’ war on cancer: Preclinical perspectives. Int J Mol Sci. 2021; 22(21): 11500. DOI: https://doi/org/10.3390/ijms222111500
CrossRef - Lu N., Malemud C. J. Extracellular signal-regulated kinase: A regulator of cell growth, inflammation, chondrocyte, and bone cell receptor-mediated gene expression. Int J Mol Sci. 2019; 20(15): 3792. DOI: https://doi.org/10.3390/ijms20153792
CrossRef - Fakhri S., Moradi S. Z., Moradi S.Y., Piri S., Varnamkhasti B.S., Piri S., Khirehgesh M.R., Bishayee A., Casarcia N., Bishayee A. Phytochemicals regulate cancer metabolism through modulation of the AMPK/PGC-1α signaling pathway. BioMed Central Cancer. 2024; 24: 1079. DOI: https://doi/org/10.1186/s12885-024-12715-7
CrossRef - Bose S., Banerjee S., Mondal A., Chakraborty U., Pumarol J., Croley C.R., Bishayee A. Targeting the JAK/STAT signaling pathway using Phytocompounds for Cancer Prevention and Therapy. Cells. 2020; 11; 9(6): 1451. DOI: https://doi/org/10.3390/cells9061451.
CrossRef - Tewari D., Priya A., Bishayee A., Bishayee A. Targeting transforming growth factor-β signalling for cancer prevention and intervention: recent advances in developing small molecules of natural origin. Clin Transl Med. 2022; 12(4): e795. DOI: https://doi/org/10.1002/ctm2.795.
CrossRef - Kaur C., Sahu S.K., Bansal K., DeLiberto L.K., Zhang J., Tewari D., Bishayee A. Targeting peroxisome proliferator-activated Receptor-β/δ, Reactive Oxygen Species and Redox Signaling with Phytocompounds for Cancer Therapy. Antioxidants Redox Signal. 2024; 41(4-6): 342-395. DOI: https://doi/org/10.089/ars.2023.0442.
CrossRef - Jit B.P., Pradhan B., Dash R., Bhuyan P.P., Behera C., Behera K.R., Sharma A., Alcaraz M., Jena M. Phytochemicals: Potential therapeutic modulators of radiation-induced signaling pathways. Antioxidants (Basel). 2021; 11(1); 19. DOI: https://doi/org/10.3390/ANTIXO11010049.
CrossRef - Zhang T., Zhang H., Shi X., Wu C., Liu B. The mechanism of resveratrol on colorectal cancer by regulating AMPK/PGC-1-alpha signaling pathway. Indian J Pharm Sci. 2023; 85(6): 1654. DOI: https://doi/org/10.36468/PHARMACEUTICAL-SCIENCES.1220
CrossRef - Rahman M.M., Sarker M.T., Alam Tumpa M.A., Yamin M., Islam T., Park M.N., Islam M.R., Rauf A., Sharma R., Cavalu S., Kim B. Exploring the recent trends in perturbing the cellular signaling pathways in cancer by natural products. Front Pharmacol. 2022; 13: 950109. DOI: https://doi.org/10.3389/FPHAR.2022.950109
CrossRef - Chirumbolo S., Bjorklund G., Lysiuk R., Vella A., Lenchyk L., Upyr T. (2018). Targeting cancer with phytochemicals via their fine-tuning of the cell survival signaling pathways. Int J Mol Sci. 2018; 19(11): 3568. DOI: https://doi.org/10.3390/IJMS19113568
CrossRef - Khan N., Jajeh F., Eberhardt E.L., Miller D.D., Albrecht D.M., Van Doorn R., Hruby M.D., Maresh M.E., Clipson L., Mukhtar H., Halberg R.B. Fisetin and 5-fluorouracil: Effective combination for PIK3CA-mutant colorectal cancer. Int J Cancer. 2019; 145(11): 3022–3032. DOI: https://doi.org/10.1002/IJC.32367
CrossRef - Khan A.W., Farooq M., Haseeb M., Choi S. Role of plant-derived active constituents in cancer treatment and their mechanisms of action. Cells 2022; 11(8): 1326. DOI: https://doi.org/10.3390/cells11081326
CrossRef - Zhou L., Zhang J., Zhao K., Chen B., Sun Z. Natural products modulating MAPK for CRC treatment: a promising strategy. Front Pharmacol. 2025; 16; 1514486. DOI: https://doi.org/10.3389/FPHAR.2025.1514486
CrossRef - Aslan A., Beyaz S., Gok O., Can M.I., Erman F., Erman O. The impact of ellagic acid on some apoptotic gene expressions: a new perspective for the regulation of pancreatic Nrf-2/NF-κB and Akt/VEGF signaling in CCl4-induced pancreas damage in rats. Immunopharmacol Immunotoxicol. 2021; 43(2): 145–152. DOI: https://doi.org/10.1080/08923973.2020.1869255
CrossRef - Bidanchi R.M., Lalrindika L., Khushboo M., Bhanushree B., Dinata R., Das M., Nisa N, Lalrinzuali S, Manikandan B., Saeed-Ahmed L., Sanjeev S., Murthy M.K., Roy V.K., Gurusubramanian G. Antioxidative, anti-inflammatory, and anti-apoptotic action of ellagic acid against lead acetate induced testicular and hepato-renal oxidative damages and pathophysiological changes in male Long Evans rats. Enviro Pollut. 2022; 302: 119048.DOI: https://doi.org/10.1016/J.ENVPOL.2022.119048
CrossRef - Gajos-Michniewicz, A., Czyz, M. Therapeutic potential of natural compounds to modulate WNT/β-Catenin signaling in cancer: Current state of art and challenges. Int J Mol Sci. 2024; 25 (23): 12804. DOI: https://doi.org/10.3390/IJMS252312804
CrossRef - Villota H., Rothlisberger S., Pedroza-Díaz J. Modulation of the canonical wnt signaling pathway by dietary polyphenols. An opportunity for colorectal cancer chemoprevention and treatment. Nutr Cancer. 2022; 74(2), 384–404. DOI: https://doi.org/10.1080/01635581.2021.1884730
CrossRef - Hasan G.M., Hassan M.I., Sohal S.S., Shamsi A., Alam M. Therapeutic targeting of regulated signaling pathways of non-small cell lung carcinoma. ACS Omega. 2023; 8(30); 26685–26698. DOI: https://doi.org/10.1021/acsomega.3c02424
CrossRef - Ge A., He Q., Zhao D., Li Y., Chen J., Deng Y., Xiang W., Fan H., Wu S., Li Y., Liu L., Wang Y. Mechanism of ferroptosis in breast cancer and research progress of natural compounds regulating ferroptosis. J Cell Mol Med. 2024; 28(1): e18044. DOI: https://doi.org/10.1111/JCMM.18044
CrossRef - Li J., Zhang J., Zhu Y., Afolabi L.O., Chen L., Feng, X. Natural compounds, optimal combination of brusatol and polydatin promote anti-tumor effect in breast cancer by targeting Nrf2 signaling pathway. Int J Mol Sci. 2023; 24(9): 8265. DOI: https://doi.org/10.3390/IJMS24098265
CrossRef - Abdalla Abdelaziz M.A., Nelson V.K., Kumarasamy M., Peraman R. Anticancer effect of polyphenolic acid enriched fractions from Grewia bracteata Roth on tumor cells and their p53 gene independent ROS mediated apoptosis in colon cancer cells. 2023; 233, 107243. DOI: https://doi.org/10.1016/J.TOXICON.2023.107243
CrossRef - Merlin J. P. J., Rupasinghe H. P. V., Dellaire G., Murphy, K. Role of dietary antioxidants in p53-mediated cancer chemoprevention and tumor suppression. Oxid Med Cell Longev. 2021(1): 9924328. DOI: https://doi.org/10.1155/2021/9924328
CrossRef - Sohel M., Biswas P., Al Amin M., Hossain M. A., Sultana H., Dey D., Aktar S., Setu A., Khan M.S., Paul P., Islam MN., Rahman MA., Kim B., Mamun A.A. Genistein, a potential phytochemical against breast cancer treatment-insight into the molecular mechanisms. 2022; 10(2): 415. DOI: https://doi.org/10.3390/PR10020415
CrossRef - Yao L., Fan Z., Han S., Sun N., Che, H. Apigenin acts as a partial agonist action at estrogen receptors in vivo. Eur J Pharmacol. 2021; 906: 174175. DOI: https://doi.org/10.1016/J.EJPHAR.2021.174175
CrossRef - Sung H., Ferlay J., Siegel R.L., Laversanne M., Soerjomataram I., Jemal A., Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: Cancer J Clin. 2021; 71: 209-249. DOI: https://doi/org/10.3322/caac.21660.
CrossRef - Siegel R.L., Miller K.D., Fuchs H.E., Jemal A. Cancer statistics, 2022. CA: A Cancer J Clin. 2022; 72: 7-33. DOI: https://doi/org/10.3322/caac.21708.
CrossRef - Sathishkumar K., Chaturvedi M., Das P., Stephen S., Mathur P. Cancer incidence estimates for 2022 & projection for 2025: Result from National Cancer Registry Programme, India. Indian J Med Res. 2022; 156 (4&5): 598-607. DOI: https://doi/org/10.4103/ijmr.ijmr_1821_22.
CrossRef - Chang J. H., Cheng C. W., Yang Y. C., Chen W. S., Hung W. Y., Chow J. M., Chen P.S., Hsiao M., Lee W.J., Chien M.H. Downregulating CD26/DPPIV by apigenin modulates the interplay between Akt and Snail/Slug signaling to restrain metastasis of lung cancer with multiple EGFR statuses. J Exp Clin Cancer Res. 2018; 37 (1): 199. DOI: https://doi/org/10.1186/s13046-018-0869-1
CrossRef - Lee W.J., Cheng T.C., Yen Y., Fang C.L., Liao Y.C., Kuo C.C., Tu S.H., Lin L.C., Chang H.W., Chen L.C,. Ho YS. Tea polyphenol epigallocatechin-3-gallate inhibits cell proliferation in a patient-derived triple-negative breast cancer xenograft mouse model via inhibition of proline-dehydrogenase-induced effects. J Food Drug Anal. 2021; 15;29(1):113-127. DOI: https://doi/org/10.38212/2224-6614.3230
CrossRef - De Lima R.M.T., Dos Reis A. C., De Menezes A.P.M., Santos J. V. O., Filho J., Ferreira J. R. O., De Alencar M.V.O.B., Da Mata A.M.O.F., Khan I.N., Islam A., Uddin S.J., Ali E.S., Islam M.T., Tripathi S., Mishra S.K., Mubarak M.S., Melo-Cavalcante A.A.C. Protective and therapeutic potential of ginger (Zingiber officinale) extract and [6]-gingerol in cancer: A comprehensive review. Phytotherap Res. 2018; 32 (10): 1885–1907.
CrossRef - Talib W.H., Awajan D., Alqudah A., Alsawwaf R., Althunibat R., Abu AlRoos M., Al Safadi A., Abu Asab S., Hadi R.W., Al Kury L.T. Targeting cancer hallmarks with epigallocatechin gallate (EGCG): Mechanistic basis and therapeutic targets. Molecules. 2024; 20;29(6):1373. DOI: https://doi/org/10.3390/molecules29061373
CrossRef - Madathil S.R., Kannappan P., Muthusami S., Muneeswari, P. Phytotherapeutics in Colorectal cancer with emphasis on target molecular signaling pathways: A review. Indian J Pharm Sci. 2022; 84(6):1358 – 1368.
CrossRef - Reynoso-Camacho R., González-Jasso E., Ferriz-Martínez R., Villalon-Corona B., Loarca-Pina G.F., Salgado L.M. Dietary supplementation of lutein reduces colon carcinogenesis in DMH-treated rats by modulating K-ras, PKB, and β-catenin proteins. Nutr Cancer. 2011; 63 (1): 39 -45. DOI: https://doi/org/10.1080/01635581.2010.516477
CrossRef - Huang J., Xu M., Fang Y.J., Lu M.S., Pan Z.Z., Huang W.Q., Chen Y.M., Zhang C,X. Association between phytosterol intake and colorectal cancer risk: a case–control study. British J Nutr. 2017; 117(6), 839–850. DOI: https://doi/org/10.1017/S0007114517000617.
CrossRef
- Wang T., Zhan R., Lu J., Zhong L., Peng X., Wang M., Tang S. Grain consumption and risk of gastric cancer. A meta-analysis. Int J Food Sci Nutr. 2020; 71: 164-175. DOI: https://doi/org/10.1080/09637486.2019.1631264
CrossRef - Xu Y., Yang J., Li K., Zhou Y. Association of whole grain, refined grain, and cereal consumption with gastric cancer risk. A meta-analysis of observational studies. Food Sci Nutr. 2018; 7: 256-265. DOI: https://doi/org/10.1002/fsn3.878
CrossRef - Liu K., Chen W., Zhou Y., Xu L., Sun X., Mao Y., Ye D. Associations between food groups and liver cancer: a systematic review and meta-analysis of observational studies. J Nutr 2023; 22(1): 30. DOI: https://doi/org/10.1186/s12937-023-00858-5
CrossRef - Reynolds A., Mann J., Cummings J., Winter N., Mete E., Te Morenga L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet. 2019; 393:434–445. DOI: https://doi/org/10.1016/S0140-6736(18)31809-9
CrossRef - Gaesser GA. Whole Grains, refined grains, and cancer risk: A systematic review of meta-analyses of observational studies. Nutrients. 2020; 12(12):3756. DOI: https://doi/org/10.3390/nu12123756
CrossRef - Wang K., Zhao J., Yang D., Sun M., Wu Y., Zhou W. Association of whole grain food consumption with lung cancer risk: A prospective cohort study. Cancer Cell Int. 2025; 25:14. DOI: https://doi/org/10.1186/s12935-025-03634-z
CrossRef - Xiao Y., Ke Y., Wu S., Huang S., Li S., Lv Z., Yeoh E.K., Lao. X., Wong S., Kim J.H., Colditz G.A., Tamimi R.M., Su X. Association between whole grain intake and breast cancer risk: A systematic review and meta-analysis of observational studies. Nutr J. 2018; 17:87. DOI: https://doi/org/10.1186/s12937-018-0394-2
CrossRef - Mahmoodi M., Mohammadi F., Rajabzadeh-dehkordi M., Jalilpiran Y., Makhtoomi M., Nouri M., Faghih S. The association between healthy diet indicator and phytochemical index with prostate cancer odds ratio: A case-control study. J Health Popul Nutr. 2024; 43:116. DOI: https://doi/org/10.1186/s41043-024-00603-6.
CrossRef - Poorojalal J., Moradi L., Mohammadi Y., Cheraghi Z., Gohari–Ensaf F. Risk factors for stomach cancer. A systematic review and meta-analysis. Epidemiol Health. 2020; 42e2020004. DOI: https://doi/org/10.4178/epih.e2020004
CrossRef - Epplein M., Zheng W., Li H., Peek R.M., Correa P., Gao J., Michel A., Pawlita M., Cai Q., Xiang Y.B., Shu X.O. Diet, Helicobacter pylori strain-specific infection, and gastric cancer risk among Chinese men. Nutr Cancer. 2014; 66(4): 550-557. DOI: https://doi/org/10.1080/01635581.2014.894096
CrossRef - Kim S.R., Kim K., Lee S.A., Kwon S.O., Lee J.K., Keum N., Park SM. Effect of red, processed, and white meat consumption on the risk of gastric cancer: An overall and dose-response meta-analysis. Nutrients. 2019; 11: 826. DOI: https://doi/org/10.03390/nu11040826
CrossRef - Farvid M.S., Sidahmed E., Spence N.D., Mante Angua K., Rosner B.A., Barnett J.B. Consumption of red meat and processed meat and cancer incidence: A systematic review and meta-analysis of prospective studies. Eur J Epidemiol. 2021; 36(9): 937-951. DOI: https://doi/org/10.1007/s10654-021-00741-9
CrossRef - Di Y., Ding L., Gao L., Huang H. Association of meat consumption with the risk of gastrointestinal cancers: a systematic review and meta-analysis. BioMed Central Cancer. 2023; 23: 782. DOI: https://doi/org/10.1186/s12885-023-11218-1
CrossRef - Bradbury, K.E., Murphy N., Key T.J. Diet and colorectal cancer in UK Biobank: A prospective study. Int J Epidemiol. 2020; 49: 246-258. DOI: https://doi/org/10.1093/ije/dyz064
CrossRef - Huang Y., Cao D., Chen Z., Chen B., Li J., Guo J. Dong Q., Liu L., Wei Q. Red and processed meat consumption and cancer outcomes. Umbrella studies. Food Chem. 2021; 356: 129697. DOI: https://doi/org/10.1016/j.foodchem.2021.129697
CrossRef - Dunneram, Y., Greenwood D.C., Cade JE. Diet and risk of breast, endometrial and ovarian cancer: UK Women’s Cohort Study. British J Nutr. 2019; 122(5): 564–574. DOI: https://doi.org/10.1017/S0007114518003665
CrossRef - Farvid M.S., Chen W.Y., Rosner B.A., Tamimi R.M., Willett W.C., Eliassen A.H. Fruit and vegetable consumption and breast cancer incidence: Repeated measures over 30 years of follow-up. Int J Cancer. 2019; 144(7): 1496–1510. DOI: https://doi.org/10.1002/ijc.31653
CrossRef - Guo X.Y., Shao X.Y., Li J.M., Li S., Li K., Li D. Fruit and vegetable intake and liver cancer risk: A meta-analysis of prospective cohort studies. Food Funct. 2019; 10(8): 4478–4485. DOI: https://doi.org/10.1039/c9fo00804g
CrossRef - Imran M., Salehi B., Sharifi-Rad J., Aslam Gondal T., Saeed F., Imran A., hahbaz M, Tsouh Fokou PV, Umair Arshad M., Khan H., Guerreiro .SG., Martins N., Estevinho L.M. Kaempferol: A key emphasis to its anticancer potential. Molecules. 2019; 19;24(12):2277. DOI: https://doi/org/10.3390/molecules24122277
CrossRef - Vieira A.R., Abar L., Vingeliene S., Chan D.S.M., Aune D., Navarro-Rosenblatt D, Stevens C, Greenwood D, Norat T.Fruits, vegetables, and lung cancer risk: A systematic review and meta-analysis. Ann Oncol. 2016; 27(1): 81–96. DOI: https://doi.org/10.1093/annonc/mdv381
CrossRef - Xu X.Y., Zhao C.N., Cao S.Y., Tang G.Y., Gan R.Y., Li H.B. Effects and mechanisms of tea for the prevention and management of cancers: An updated review. Crit Rev Food Sci Nutr. 2020; 60(10):1693-1705. DOI: https://doi/org/10.1080/10408398.2019.1588223
CrossRef - Li C., Xu Y., Zhang J., Zhang Y., He W., Ju J., Wu Y, Wang Y. The effect of resveratrol, curcumin, and quercetin combination on immuno-suppression of tumor microenvironment for breast tumor-bearing mice. Sci Rep. 2023; 16;13(1):13278. DOI: https://doi.org/10.1038/s41598-023-39279-z
CrossRef - Wu Z.Y., Chen J.L., Li H., Su K., Han Y.W. Different types of fruit intake and colorectal cancer risk: A meta-analysis of observational studies. World J Gastroenterol. 2023; 29 (17): 2679-2700. DOI: https://doi/org/10.3748/wjg.v29.i17.2679
CrossRef - Balali A., Askari G., Anjom-Shoae, J., Sadeghi, O. Association between nut consumption and prostate cancer risk in adults: A systematic review and dose-response meta-analysis of observational studies. Nutr, Metab Cardiovas Dis. 2023; 33(7): 1293–1307. DOI: https://doi.org/10.1016/j.numecd.2023.04.004
CrossRef - Hong M.Y., Moore J., Nakagawa A., Nungaray V. Effects of Mixed Nuts on Colonic Cell Proliferation and Ptgs2and Rela Gene Expression. Anticancer Res. 2022; 42(9):4285-4292. DOI: https://doi/org/10.21873/anticanres.15928
CrossRef - Jin S., Je, Y. Nuts and legumes consumption and risk of colorectal cancer: A systematic review and meta-analysis. Eur J Epidemiol. 2022; 37(6), 569–585. DOI: https://doi.org/10.1007/s10654-022-00881-6
CrossRef - Huang Y., Chen Q., Liu Y., Tian R., Yin X., Hao Y., Yang Y., Yang J., Li Z., Yu S., Li H., Wang G. Association between tea consumption and colorectal cancer: A systematic review and meta-analysis of a population-based study. BioMed Central Gastroenterol. 2023; 23(1):294. DOI: https://doi/10.1186/s12876-023-02928-8
CrossRef - Hu Y., Lin Q., Zhao H., Li X., Sang S., McClements D.L. et al. Bioaccessibility and bioavailability of phytochemicals. Influencing factors, improvement, and evaluations. Food Hydrocoll. 2023; 108165. DOI: https://doi/org/10.1016/j.foodhyd.2022.108165
CrossRef - Mishra A.K., Singh R., Rawat H., Kumar V., Japtap C., Jain A. The influence of food matrix on the stability and bioavailability of phytochemicals. A comprehensive review. Food Humanity. 2024; 100202. DOI: https://doi/org/10.1016/j.foohum.2023.12.010
CrossRef - Shi M., Gu J., Wu H., Rauf A., Emran T.B., Khan Z. et al. Phytochemicals, nutrition, metabolism, bioavailability, and health benefits in lettuce. A comprehensive review. 2022; 11:1158. DOI: https://doi.org/10.3390/antiox11061158
CrossRef - Nicolescu A., Babota M., Barros L., Rocchetti G.., Lucini L., Tanase C. Mocan A., Bunea C.I., Crișan G. Bioaccessibility and bioactive potential of different phytochemical classes from nutraceuticals and functional foods. Front Nutr. 2023; 27(10): 1184535. DOI: https://doi/org/10.3389/fnut.2023.1184535
CrossRef - Dabeek W.M., Marra M.V., Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients. 2019; 11(10): 2288. DOI: https://doi/org/10.3390/nu11102288
CrossRef - Kaushal N., Singh M., Singh S.R. Flavonoids: Food associations, therapeutic mechanisms, metabolism and nanoformulations. Food Res Int. 2022; 157: 111442. DOI: https://doi/org/10.1016/j.foodres.2022.111442
CrossRef - Li D., Sun S., Chen J. Research progress on classification, source, application of phytosterol esters, and their thermal oxidation stability. Grain Oil Sci Technol. 2024; 7(1): 1-11. DOI: https://doi/org/10.1016/j.gaost.2023.12.003
CrossRef - Pokorski P., He R., Kurek M. A. Advancing protein hydrolysis and phytosterol encapsulation: Emerging trends and innovations in protein-based microencapsulation techniques – A comprehensive review. Food Res Int. 2024; 196: 115012. DOI: https://doi/org/10.1016/j.foodres.2024.115012
CrossRef - Sahraeian S., Rashidinejad A., Golmakani M.T. Recent advances in the conjugation approaches for enhancing the bioavailability of polyphenols. Food Hydrocoll. 2024; 146: 109221. DOI: https://doi/org/10.1016/j.foodhyd.2023.109221
CrossRef - Zhou H., Zheng B., Mc Clements D.J. Encapsulation of lipophilic polyphenols in plant-based nanoemulsions: impact of carrier oil on lipid digestion and curcumin, resveratrol and quercetin bioaccessibility. Food Funct. 2021; 12: 3420-3432.
CrossRef - Jiang X., Zuo L., Gao S., Yang Q., Li Y., Chen Y., Xie X., Peng C. Green production pathways, instability, and stability of resveratrol: A systematic review. J Food Biochem. 2025; 8210896. DOI: https://doi/org/10.1155/jfbc/8210896
CrossRef - Gaber A.A.A., Nasr M., Hamzawy M.A. Pterostilbene lipid nanocapsules as a promising system for treatment of hepatocellular carcinoma: Formulation, characterization, and cellular/in vivo appraisal. J Drug Deliv Sci Technol. 2023; 92(4): 105310. DOI: https://doi/org/10.1016/j.jddst.2023.105310
CrossRef - Molteni C., La Motta C., Valoppi F. Improving the bioaccessibility and bioavailability of carotenoids by means of nanostructured delivery systems: A comprehensive review. Antioxidants. 2022; 28:11(10): 1931.DOI: https://doi/org/10.3390/antiox11101931
CrossRef - Liu M., Wang F., Pu C., Tang W., Sun Q. Nanoencapsulation of lutein within lipid-based delivery systems: Characterization and comparison of zein peptide stabilized nano-emulsion, solid lipid nanoparticle, and nano-structured lipid carrier. Food Chem. 2021; 129840. DOI: https://doi/org/10.1016/j.foodchem.2021.129840
CrossRef
List of Abbreviations
17HSD 17β: 17β-hydroxysteroid dehydrogenases
ATA: Antcin – A
BAX: BCL2 Associated X
BCL-2: B cell lymphoma 2
BT – 20: Breast cancer cell line – 20
BT – 549: Breast cancer cell line – 549
cagA: Cytotoxin–associated gene A
CAMP: Campesterol
CAT: Catalase
CCND1: Cyclin D1
CCS: Case-control studies
CDK: Cyclin-dependent kinases
CFL2: Cofilin-2
COX: Cyclooxygenase
CS: Cohort study
DMH: Dimethylhydrazine
DNMT1: DNA methyltransferase 1
EEP: Ethanolic extract of propolis
EGCG: Epigallocatechin gallate
EMT: Epithelial-mesenchymal transition
ERGO: Ergosterol
ERK: Extracellular signal-regulated kinase
FADD: Fas-associated protein with death domain
FOXO: Forkhead box transcription factors
G2/M: Second gap phase/Mitosis
GPX: Glutathione peroxidase
GRP78: Glucose-regulated protein 78
HCA: Heterocyclic amines
HEK293T: Human embryonic kidney 293 cells
HER2: Human epidermal growth factor receptor 2
HIF 1α: Hypoxia–inducible factor 1 alpha
hnRNPA1: Heterogeneous nuclear ribonuclear protein
H. pylori: Helicobacter pylori
HUVEC: Human umbilical vein endothelial cells
IL-6: Interleukin
JAK/STAT: Janus kinases/Signal transducer and activator of transcription
LEF -1: Lymphoid enhancer factor
MAPK: Mitogen–activated protein kinase
MBE: Muscadine grape berry extract
MCF7: Michigan Cancer Foundation 7
MDA – MB – 231: MD Anderson – metastatic breast 231
MDA – MB – 436: MD Anderson – metastatic breast 436
MEK: Mitogen–activated extracellular signal-regulated kinase
MIC 1: Macrophage inhibitory cytokine – 1
MMP: Matrix metalloproteinase
MTA 1: Metastasis-associated protein 1
MTase: Methyltransferase
mTOR: Mammalian target of rapamycin
NCD: Non-communicable diseases
NFR2: Nuclear factor erythroid 2-related factor 2
NLRP: Nucleotide-binding and leucine-rich repeat protein
NSCLC: Non-small cell lung cancer
OR: Odds ratio
P13K/AKT: phosphatidylinositol 3 kinase/Akt kinase
p53: Tumour protein
PAI-1: Plasminogen activator inhibitor-1
PAH: Polycyclic aromatic hydrocarbons
PCNA: Proliferating cell nuclear antigen
PG: Prostaglandin
PIK3CA: Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha
PPARS: Peroxisome proliferator-activated receptors
PTEN: Phosphatase and tensin homolog
Raf: Rapidly accelerated fibrosarcoma
Ras: Rat sarcoma
RASSFIA gene: Ras association domain family member 1
ROS: Reactive oxygen species
RR: Relative risk
SITO: Sitosterol
SOD: Superoxide dismutase
STAT3: Signal transducer and activator of transcription 3
STIG: Stigmasterol
TGF-β: Transforming growth factor β
TIMP: Tissue inhibitors of metalloproteinases
TLR: Toll-like receptors
TNBC: Triple negative breast cancer
TNF–α: Tumor necrosis factor-alpha
TRAIL: Tumor Necrosis Factor (TNF)-related apoptosis-inducing ligand
VEGF: Vascular endothelial growth factor

This work is licensed under a Creative Commons Attribution 4.0 International License.






