A Potential Role of Xenometabolites as Nutraceutical: A Concurrent Review
 , Ali Imran2, , 3, Muhammad Umair Arshad2, Iqra Aslam2, Mohd Asif Shah4, Umber Shehzadi2, Ifrah Usman2, , Safura Aslam2
, Ali Imran2, , 3, Muhammad Umair Arshad2, Iqra Aslam2, Mohd Asif Shah4, Umber Shehzadi2, Ifrah Usman2, , Safura Aslam2 1Department of Food and Nutrition, Acharya Prafulla Chandra College, Kolkata-India.
2Department of Food Science, Government College University Faisalabad-Pakistan.
3Department of Clinical Nutrition, NUR International University, Lahore, Pakistan.
4Adjunct Faculty, University Center for Research and Development, Chandigarh University, Gharuan, Mohali India.
Corresponding Author E-mail: sunandafnt@gmail.com
DOI : http://dx.doi.org/10.12944/CRNFSJ.12.1.02
Download this article as:
![]()
The gut microbiota has the capacity to de-novo manufacture or change endogenous and exogenous substances to produce or alter xenometabolites (i.e., non-host-derived metabolites). A wide-scale characterization of these metabolites is still lacking, despite rare instances of xenometabolites impacting host health and illness. Numerous studies have been conducted to investigate how the gut microbiome affects individual function and health, including links between specific intestinal microorganism populations and metabolites and the health of the systemic-immune system and gastrointestinal tract. The current review article delves into the sources of xenometabolites and the role of modeling in addressing the complexity of the xenometabolites process, as well as various nutraceutical benefits such as antibiotics, anti-tumor, and anti-cancer action.
KEYWORDS:Antibiotics; Anti-cancer; Anti-tumor; Promote growth; Xenometabolites
Introduction
Although orally ingested xenometabolites (small compounds originating from natural products, foods, medicines, and industrial origins) have been significantly discovered, microbe-generated xenometabolites have received less attention. It is becoming increasingly clear that the intestinal flora impacts host health and illness1. The role of microbially formed xenometabolites, or bacterial transformation of moderator metabolites, to moderator physiology is well substantiated by the available information2. It has been revealed that colonic epithelial cells’ synthesis of butyrate stimulates -oxidation and promotes the proliferation of intestinal regulatory T cells, establishing a positive response loop that keeps the intestinal epithelia and obligatory anaerobes in a mutually favorable relationship3. On top of the straight effects on intestinal tissues, several reports have recommended a link between microbe-derived xenometabolites and heart disease4 and several other diseases5. Metabolomics has been a primary tool to recognize target tissues for some microbe-derived xenometabolites in various host tissues6. However, on-guard evaluation of assumed xenometabolite formation, absorption, shipment, and interorgan flux has never been conducted. The objective of the recent study was to characterize the roots and potential organ objectives of alleged xenometabolites and microbe-modified metabolic products in good shape post-absorptive awake pigs using a specialized metabolomics framework (XenoScan) nourished in xenometabolites, their variants, and gastrointestinal (GI)-related substances (bile acids). Following investigation employed the XenoScan living up on the library of real standards from the Arkansas Children’s Nutrition Center and centered on known and speculative xenometabolites and their variants7. Maximum range were produced for all the metabolites in our internal collection (i.e., unrestricted to only microbial commodified metabolites and xenometabolites). The biggest outer layer of microbial contact with the human defense system is made up by intestinal commensals, or the gut microbiota, which outnumber human body cells. This dynamic interface, which is made up of several metabolic, immune, and provocative responses, survive in a precarious stability between disease and symbiosis and could affect a variety of physiological features of the host. High-throughput arrangement, metabolite characterization, and bioinformatics data are modifying our information of the relationships of the stomach microbiota remoter, despite accumulating confirmation that the digestive system microbiome has a significant part in carcinogenesis8. The investigation of the footprint of microbiota on medication placement, undertaking, and dreadfulness, or “pharmacomicrobiomics,” in particular, is highlighting the vital role that microbes play in the effectiveness and toxic effect of treatment for cancer9. By modifying host-chemotherapeutic-microbiota interplay and personalizing chemotherapy procedure depending on an assessment of the bacterial habitat of a person (the genetic makeup of their microorganisms), pharmacomicrobiomics has the potential to improve curative order and eliminate harmful effects10. A deeper comprehension of the tangled interactions between the immune system and the gut bacteria may set about the evolution of a new chemotherapeutic plan of action11.
Xenometabolites
It is believed that xenometabolites derived from plant and microbial sources have both helpful and harmful impacts on host physiology. There are few studies examining xenometabolite tissue uptake and absorption. Kelly et al.12 assessed the interorgan flow of elucidated familiar and putative xenometabolites, derivative products, and blistering acids in a mindful catheterized pig model. It has been demonstrated that the bacteria in the digestive tract may alter or metabolize various xenobiotics, including modern cancer medications, millennia-old analgesics, and food supplements13. Recent investigation has also revealed the viability of utilizing and modifying such a microbe-mediated xenometabolism to enhance host health or prevent adverse drug reactions. For instance, Wallace et al.14 shown that blocking bacterial b-glucuronidase can prevent a harmful bioconversion of the cancer medication irinotecan. Probiotic bacteria, such as Lactobacillus sp., have been demonstrated to alleviate diarrhea, illnesses linked to Clostridium difficile, and other antibiotic adverse effects15. Although few workrooms have devoted genuine quality athenaeum that cover the broad range of substances involved in metabolism caused by bacterial activity, metabolomics has proven highly useful in identifying molecular assay of microbial metabolism. Identification of microbial-specific metabolites is further complicated by the fact that the majority of large-scale metabolomics instruments use basic and spectroscopic libraries that often include human and animal metabolic pathways. To solve this problem, a metabolomics manifesto was created that allows for a stronger focal point on “nonhost” xenometabolites, xenometabolite variants, cometabolites, & congeners. This platform may be used to better analyze changes in the microbial environment. Because of developments in omics technology, the molecular routes of xenometabolism inside the intestinal microbiota have now begun to emerge through the recognition of the microbes and enzymes in charge16. Today, a significant fragment of the bacterial species colonising the human gut can be identified with the aid of metagenomics tools17. Additionally, these tools are illuminating the gut microbiome’s genetic repertoire in an unprecedented level of detail.
Sources of xenometabolites
The primary biopolymeric component of plants, starch, which includes kernels, nut, and radix, appears in distinctive sandy forms in a variety of sizes and structures18-19. The main medium of physiological energy found in the human food is starch, which is why it is generally categorized as a readily accessible carbohydrate. Nutritionists & food technologists especially have been interested in the importance of starch as a functional polysaccharide. Both the food and non-food sectors use starch in a wide range of products. In addition to providing calories, starch is also found major part in the texture of food, which affects its organoleptic qualities. This is true of most processed food systems. Dietary fibre (DF) is basically made up of outer covering of plant constituents like polysaccharides and lignin and is commonly described as the large molecules in the food that withstand breakdown by human inner enzymes. In the past 20 years, DF has significantly lowered the risk of a wide range of illnesses, including diabetes mellitus, heart conditions, cancer of large intestine, constipation, and diverticulosis. High-fibre diets have slowed nutrient uptake, increased fecal volume, decreased blood lipids, reduced risk of cancer of the large intestine, an obstacle to breakdown, mobile intestinal parts, longer fecal elimination times, and properties that are fermentable. The multiple positive physiological impacts of DF and how they relate to how the large and small intestines operate20. Dietary fibre is a mixture of structurally diverse components such cellulose, hemicelluloses, pectic substances, gums, mucilages, complex carbohydrates, other polysaccharides, and lignin, a non-carbohydrate polymer made of phenyl propane remainings, rather than making up a specific chemical group. DF is primarily divided into two further forms: soluble DF and insoluble DF. Most research on the interactions among the bacteria of digestive tract and the host has concentrated on relations among the fullness of particular microbes and particular (patho) physiological variables, allowing for the formation of hypotheses concerning the possible bacterial taxa that may be tangled. When combined with the vast biomass of microorganisms in the intestinal system and the complex chemical mixes in food, the gut microbiome has a great genetic makeup (number of gene variants and genes) that has a significant nonenzymatic and enzymatic capacity for producing xenometabolites. De novo formation, bacterial transformation of external parts (such as phytochemicals, fibres, oligosaccharides, pharmaceutical compounds, etc.), and bacterial alternation of molecules originating from hosts can all be used to achieve this (central bile acids, urea, etc.). Due to the fact that both the host and the microbe are involved in their metabolism, the latter are also known as “cometabolites” (s)21-22.
Role of modeling in tackling complexity of xenometabolic processes
Because of three key factors—the broad license of metabolic enzymes, the constructional diversity of the bacteria presents in digestive track, as well as the interconnections among the recipient and the microbe-mediated xenometabolism—xenometabolic activities in the gut can be extremely complicated. According to Tawfik23, the versatility of metabolic enzymes increases the number of potential pathways by which a xenobiotic chemical could be digested or changed when combined with the microbiota’s enzymatic reserve. This complexity is further increased by the host-microbiota interactivity during the enterohepatic cycle, the textural variety and natural variability of the microbiota, and other factors. The appeal for modeling and simulation of xenometabolism is increased by these inherent complexities and the limitations in collecting in vivo studies from human participants. A comprehensive modeling framework that considers coupled host-microbiota xenometabolism might be useful for developing drugs and developing individualised medication plans. A platform like this would make it possible to forecast probable xenometabolic pathways and so create provable hypotheses. In spite of the fact that no manifesto now addresses all of the many difficulties during modeling xenometabolism, there are a number of technologies that can handle the crucial individual phases. These methods include modeling biochemical interactions in microbial populations and forecasting enzyme-level bioconversion24-25. Here, discuss these various tools and outline the key ideas that guide how they operate and also provide a number of xenometabolism examples and talk about how computational techniques that focus on various xenometabolic processes may be merged into a unified platform.
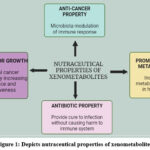 |
Figure 1: Depicts nutraceutical properties of xenometabolites. |
Anti-cancerous property
Chemotherapeutic response is significantly influenced by the microbiota’s modulation of the immune response. It has been demonstrated that continual communication between the immune system’s innate and adaptive responses and gut bacteria at the mucosal surface controls inflammation and sets the immunological tone26. Chemotherapeutic drugs may cause mucosal epithelial destruction, which may lead to bacterial translocation. Greater exposure to harmful microorganisms may result in systemic infection, but it can also stimulate the adaptive immune responses, enhancing the host’s response to chemotherapy27. There are several different ways that immunomodulation can happen. Bacterial translocation, T-helper 17 cell activation, intraluminal myeloid cell activation, and microbiota-driven T-cell activation are crucial in aiding new anti-cancer immunotherapy28. Cyclophosphamide is made more effective by these factors as well as by oxaliplatin.
Anti-tumour growth
Sivan et al.29 found that genetically comparable mice from two parties Farms (Tac) and Jackson Laboratory—grew melanoma differently (Jax). When compared to Jax mice, Tac animals had more aggressive tumor growth, which could be reversed by co-housing or colonic transfer by Jax mice. The reverse fecal transfer did not result in accelerated tumor development. Additionally, Tac mice had a reduced ability to respond to the chemotherapy drug anti-programmed necrosis 1 ligand-1 monoclonal antibodies (anti-PD-L1, pembrolizumab) as compared to Jax animals. The existence of Bifidobacterium spp. in the intestinal flora of Jax mice was found to be responsible for the reduced basal cancer progression and improved anti-cancer response to anti-PD-L1. These bacteria are thought to cooperate with dendritic cells to stimulate T cells and enhance a shielding anti-cancer response. Bifidobacterium spp. management to Tac mice was able to delay the formation of basal tumors to improve the anti-tumor impact of anti-PD-L129. In addition to promoting intestinal flora and anti-tumor responses, the gut microbiota also induces chronic dysregulated inflammation, which has genotoxic consequences and can cause carcinogenesis30. The makeup of the gut bacteria and the equilibrium between hazardous microbial population within the microbiome ultimately determine if the gut bacteria will promote colon cancer or create a healthy symbiotic connection with the host. It is unknown, nevertheless, if this “dysbiosis” might precede or lead to colorectal cancer (CRC). Studies on mice overwhelmingly support the idea that the gut microbiota may influence CRC susceptibility and may be used as early diagnostic indicators as well as treatment targets. Such clinical studies must to take into account the impact of each person’s race, way of life, food, material type, the position of the tissue collected, and the gut microbiome. There have been a number of significant changes in the taxonomic levels in the gut bacteria ecology of CRC patients. In both the intestinal mucosa and the patients’ feces, there is a rise in Bacteroides, Fusobacteria, as well as Proteobacteria and a reduction in Firmicutes31. The three best known variants in colorectal carcinogenesis are enterotoxigenic strains of B. fragilis, polyketide synthase (PKS) positive variants of E. coli, and Fusobacterium nucleatum32. The intestinal bacteria are a zestful shape that is affected by complicated interlinkage among several variables, including host immunity, chemotherapeutics, concurrent medicines, environment, and food during cancer treatment11. Both therapeutic advantages and disadvantages might result from the microbiota’s functional variety. It will need a committed and cutting-edge systems medicine approach to gain a thorough understanding of the purpose of the intestinal microbiota, including their role in chemotherapy. It is extremely difficult to manipulate the gut microbiota in a way that maximizes chemotherapy effectiveness while minimizing negative effects33.
Table 1: Nutraceutical properties of xenometabolite
|
Metabolite process |
Nutraceutical properties |
Study |
Reference |
|
Xenometabolite (Complex biochemical process) |
modeling of the intestinal flora liver biological change |
study of microbes and enzymes |
(34) |
|
Xenometabolite Millennia |
determining chemotherapeutic (cancerous) lung cancer |
Human (study was conducted in 15 centers in six countries of Central and Eastern Europe) |
(35, 36) |
|
Xenometabolite hubs |
UC Davis Type 2 Diabetes Mellitus (UCD–T2DM) |
Male rats |
(7) |
|
Xenometabolites dietary fiber |
Detoxification Organs Improve liver (alter liver metabolism) and kidney (reduced indole cresol burden) function |
animal models |
(37) |
Antibiotic property
Numerous studies have examined (see Table 1) the links between antibiotic usage and chemotherapy effectiveness, but few have addressed the clear confirmation that microbiota dysbiosis brought on by antibiotic treatment increases the possibility of cancer. In breast cancer (BC) patients having febrile neutropenia (FN), a serious side effect brought on by chemotherapy, antibiotics are needed to prevent and cure harmful bacterial infections38. Different infections can be successfully treated with antibiotics. Intestinal dysbacteriosis results, however, since their use cannot be pinpointed as accurately as targeted medications. A growing body of evidence indicates that the gut microbiota affects the effectiveness of anti-tumor treatment, such as chemotherapy, immunotherapy, radiation treatment, and surgery, used to treat complex tumors (melanoma, lung disease, and colorectal cancer). The gut microbiota plays a role in many aspects of human physiological functions, from generating nutrients and vitamins to combating pathogenic organisms and defending immune response advancement and outer covering mucosa equilibrium39.
Contrary to specific pathogen-free (SPF) mice, antibiotic-tested mice exhibited oxaliplatin (OXA) chemoresistance in colon cancer and lymphoma, suggesting that antibiotic exposure was linked to decreased chemotherapy effectiveness40. The link between the impact of neoadjuvant treatment and antibiotics on BC requires more investigation. In BC patients with FN, a serious side effect of chemotherapy, antibiotics are needed to avert and cure infectious bacterial infections38. The use of antibiotics may be linked to decreased neoadjuvant treatment effectiveness and a worse outcome in BC patients. The current study tried to assess how antibiotic administration affects the effectiveness and prognosis of neoadjuvant treatment in BC patients. Results anticipate serving as a foundation for future treatment ideas for BC patients who need antibiotics during neoadjuvant therapy41. The cells that have xenometabolic competence identical to that which happens in vivo are the best at predicting human toxicity. This characteristic has a drawback in that detoxifying metabolism may cause cytotoxicity to be underestimated
Promotion of metabolism
Metabolism is the process of generating energy by the breakdown of large molecules for the creation of desired molecules, large molecules include fats, carbohydrates and proteins, paired by minerals and vitamins as well. cytochrome P450 oxidases, UDP-glucuronosyltransferases, and glutathione ”S”-transferases metabolise xenometabolites in human beings. There are three phases of enzyme action: prime step is to oxidize the xenobiotic (phase I) and then attach water-soluble groups onto the molecule (phase II). The molecules from phase II that are water-soluble are then pumped out of cells and in multicellular organisms may be further metabolized before being excreted (phase III). Novel plasma metabolite work action reflective of enhanced metabolic health (insulin sensitivity, fitness, reduced body weight) were caught into sight before and after a 14–17week weight loss and exercise intervention in sedentary, obese insulin-resistant women. To have check and balance on potential visibal effects of diet- or microbiome-derived molecules on the systemic metabolome, sampling was during a tightly-controlled feeding test week paradigm. Intervention- and insulin-sensitivity associated were revealed by Pairwise and multivariate analysis: (1) Changes in plasma xeno-metabolites (“non-self” metabolites of dietary or intestinal microbial origin) following an oral glucose tolerance test (e.g., higher post-OGTT propane-1,2,3-tricarboxylate [tricarballylic acid]) or in the overnight-fasted state (e.g., lower γ-tocopherol)42; Enhanced indices of saturated of appreciably-long chain fatty acid elongation capacity43.
Conclusion
The gut microbiota is rapidly becoming recognized as an important location of pharmaceutical and other xenobiotic chemical metabolism important to human health. Although orally ingested xenometabolites (small compounds originating from natural products, foods, medicines, and industrial origins) have been significantly discovered, microbe-generated xenometabolites have received less attention. A growing body of evidence indicates that the gut microbiota affects the effectiveness of anti-tumor treatment, such as chemotherapy, immunotherapy, radiation treatment, and surgery, used to treat hard tumours (melanoma, lung disease, and colorectal cancer). So, in conclusion xenometabolites helps to boost the host immune system.
Acknowledgement
Authors are thankful Government College University for providing literature collection facilities. We confirm the final authorship for this manuscript, and we ensure that anyone else who contributed to the manuscript but does not qualify for authorship has been acknowledged with their permission. We acknowledge that all listed authors have made a significant scientific contribution to the research in the manuscript approved its claims and agreed to be an author.
Funding Sources
The authors declare that no funds, grants, or other support were received during the preparation of this manuscript.
Conflict of Interest
Authors declare that they have no conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal, subjects, or any material that requires ethical approval
References
- Lynch S.V., Pedersen O. The human intestinal microbiome in health and disease. New England Journal of Medicine. 2016;375: 2369-2379.
CrossRef - Vatanen T., Kostic A.D., d’Hennezel E., Siljander H., Franzosa E.A., Yassour M., Kolde R., Vlamakis H., Arthur T.D., Hämäläinen A.-M. Variation in microbiome LPS immunogenicity contributes to autoimmunity in humans. Cell. 2016;165: 842-853.
CrossRef - Byndloss M.X., Olsan E.E., Rivera-Chávez F., Tiffany C.R., Cevallos S.A., Lokken K.L., Torres T.P., Byndloss A.J., Faber F., Gao Y. Microbiota-activated PPAR-γ signaling inhibits dysbiotic Enterobacteriaceae expansion. Science. 2019;357: 570-575.
CrossRef - Senthong V., Li X.S., Hudec T., Coughlin J., Wu Y., Levison B., Wang Z., Hazen S.L., Tang W.W. Plasma trimethylamine N-oxide, a gut microbe–generated phosphatidylcholine metabolite, is associated with atherosclerotic burden. Journal of the American College of Cardiology. 2016;67: 2620-2628.
CrossRef - Yoshimoto S., Loo T.M., Atarashi K., Kanda H., Sato S., Oyadomari S., Iwakura Y., Oshima, K., Morita H., Hattori M. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature. 2013; 499:97-101.
CrossRef - Ruebel M.L., Piccolo B.D., Mercer K.E., Pack L., Moutos D., Shankar K., Andres A. 2019. Obesity leads to distinct metabolomic signatures in follicular fluid of women undergoing in vitro fertilization. American Journal of Physiology-Endocrinology and Metabolism. 2019;316: E383-E396.
CrossRef - Mercer K.E., Yeruva L., Pack L., Graham J.L., Stanhope K.L., Chintapalli S.V., Wankhade U.D., Shankar K., Havel P.J., Adams S.H. Xenometabolite signatures in the UC Davis type 2 diabetes mellitus rat model revealed using a metabolomics platform enriched with microbe-derived metabolites. American Journal of Physiology-Gastrointestinal and Liver Physiology.2020;319: G157-G169.
CrossRef - Schwabe R. F., Jobin, C. The microbiome and cancer. Nature Reviews Cancer. 2013;13(11):800-812.
CrossRef - Rizkallah M. R., Saad R., Aziz R. K. The Human Microbiome Project, personalized medicine and the birth of pharmacomicrobiomics. Current Pharmacogenomics and Personalized Medicine (Formerly Current Pharmacogenomics). 2010; 8(3):182- 195
CrossRef - Vétizou M., Pitt J.M., Daillère R., Lepage P., Waldschmitt N., Flament C., Rusakiewicz S., Routy B., Roberti M.P., Duong C.P., et al: Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015;350:1079-1084.
CrossRef - Alexander J. L., Wilson I.D., Teare J., Marchesi J.R., Nicholson J.K., Kinross J.M. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat Rev Gastroenterol Hepatol. 2017 Jun;14(6):356-365.
CrossRef - Kelly E. Mercer, Gabriella A. M. Ten Have, Lindsay Pack, Renny Lan, Nicolaas E. P. Deutz, Sean H. Adams, and Brian D. Piccolo Net release and uptake of xenometabolites across intestinal, hepatic, muscle, and renal tissue beds in healthy conscious pig. Am J Physiol Gastrointest Liver Physiol. 2020 Aug 1;319(2): G133-G141.
CrossRef - Zheng X., Zhao A., Xie G., Chi Y., Zhao L., Li,H., … & Jia, W. Melamine-induced renal toxicity is mediated by the gut microbiota. Science translational medicine. 2013;5(172):172ra22-172ra22.
CrossRef - Wallace B. D., Wang H., Lane K. T., Scott J. E., Orans J., Koo J. S., … & Redinbo M. R. Alleviating cancer drug toxicity by inhibiting a bacterial enzyme. Science. 2010;330(6005):831-835.
CrossRef - Hickson M. Probiotics in the prevention of antibiotic-associated diarrhoea and Clostridium difficile infection. Therapeutic advances in gastroenterology. 2011; 4(3) :185-197.
CrossRef - Haiser H. J., Gootenberg D. B., Chatman K., Sirasani G., Balskus E. P., Turnbaugh P. J. Predicting and manipulating cardiac drug inactivation by the human gut bacterium Eggerthella lenta. Science. 2013;341(6143) :.295-298.
CrossRef - The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature. 2012; 486:207–214.
CrossRef - Tharanathan R. N. Starch: The polysaccharide of high abundance and usefulness. Journal of Scientific and Industrial Research. 1995;54(8):452-458.
- Tharanathan R. N., Mahadevamma S. Grain legumes—a boon to human nutrition. Trends in Food Science & Technology. 2003;14(12): 507-518.
CrossRef - Turner JR. Intestinal mucosal barrier function in health and disease. Nature reviews. Immunology. 2009;9(11):799–809.
CrossRef - Libert R., Van Hoof F., Laus G., De Nayer P., Jiwan, J. L. H., De Hoffmann, E., & Schanck, A. Identification of ethyl succinyl carnitine present in some human urines. Clinica chimica acta. 2005; 355(1-2): 145-151.
CrossRef - Yang S., Minkler P., Hoppel C. cis-3, 4-Methylene-heptanoylcarnitine: characterization and verification of the C8: 1 acylcarnitine in human urine. Journal of Chromatography B. 2007;857(2) :251-258.
CrossRef - Tawfik O. K. A. D. S. Enzyme promiscuity: a mechanistic and evolutionary perspective. Annual review of biochemistry.2010 ;79:471-505.
CrossRef - Gao J., Ellis L. B., Wackett L. P. The University of Minnesota Pathway Prediction System: multi-level prediction and visualization. Nucleic acids research. 2011; 39(suppl_2): W406-W411.
CrossRef - Thiele I., Heinken A., Fleming R. M. A systems biology approach to studying the role of microbes in human health. Current opinion in biotechnology. 2013; 24(1) :4-12.
CrossRef - Perez-Chanona E., Trinchieri G. The role of microbiota in cancer therapy. Current opinion in immunology. 2016;39 :75-81.
CrossRef - Erdman S. E., Poutahidis T. Gut microbiota modulate host immune cells in cancer development and growth. Free Radical Biology and Medicine. 2017;105: 28-34.
CrossRef - Daillère R., Vétizou M., Waldschmitt N., Yamazaki T., Isnard C., Poirier-Colame V., … & Zitvogel L. Enterococcus hirae and Barnesiella intestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity. 2016; 45(4) :931-943.
CrossRef - Sivan A., Corrales L., Hubert N., Williams J.B., Aquino-Michaels K., Earley Z. M., Benyamin F.W., Lei Y.M., Jabri B., Alegre M.L., Chang E.B., Gajewski T.F. Commensal Bifidobacterium promotes anti-tumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015 Nov 27;350(6264):1084-9.
CrossRef - Park C. H., Eun C. S., Han D. S. Intestinal microbiota, chronic inflammation, and colorectal cancer. Intestinal research. 2018; 16(3) :338.
CrossRef - Yu L. C. H., Wei S. C., Ni, Y. H. Impact of microbiota in colorectal carcinogenesis: lessons from experimental models. Intestinal research. 2018 Jul;16(3):346-357.
CrossRef - Feng Q., Liang S., Jia H., Stadlmayr A., Tang L., Lan Z., … & Wang J.Gut microbiome development along the colorectal adenoma–carcinoma sequence. Nature communications. 2015; 6(1):1-13.
CrossRef - Shen S., Lim G., You Z., Ding W., Huang P., Ran C., … & Mao, J. Gut microbiota is critical for the induction of chemotherapy-induced pain. Nature neuroscience. 2017;20(9):1213-1216.
CrossRef - Klünemann M., Schmid M., Patil K. R. Computational tools for modeling xenometabolism of the human gut microbiota. Trends in biotechnology.2014; 32(3) :157-165.
CrossRef - Wang Z., Klipfell E., Bennett B. J., Koeth R., Levison B. S., DuGar B., … & Hazen S. L. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472(7341):57-63.
CrossRef - McKay J. D., Hashibe M., Hung R. J., Wakefield J., Gaborieau V., Szeszenia-Dabrowska N., … & Brennan P. Sequence variants of NAT1 and NAT2 and other xenometabolic genes and risk of lung and aerodigestive tract cancers in Central Europe. Cancer Epidemiology Biomarkers & Prevention. 2008;17(1):41-147.
CrossRef - Kieffer D. A., Martin R. J., Adams S. H. Impact of dietary fibers on nutrient management and detoxification organs: gut, liver, and kidneys. Advances in Nutrition. 2016; 7(6) :1111-1121.
CrossRef - Hashiguchi Y., Kasai M., Fukuda T., Ichimura T., Yasui T., Sumi, T. Chemotherapy-induced neutropenia and febrile neutropenia in patients with gynecologic malignancy. Anti-Cancer Drugs. 2015 Nov;26(10):1054-60.
CrossRef - Villéger R., Lopès A., Carrier G., Veziant J., Billard E., Barnich N., Gagnière J., Vazeille E., Bonnet M. Intestinal microbiota: a novel target to improve anti-tumor treatment?. International journal of molecular sciences. 2019 Sep 17;20(18):4584.
CrossRef - Iida N., Dzutsev A., Stewart C. A., Smith L., Bouladoux N., Weingarten R. A., Goldszmid, R. S. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013;342(6161) :967-970.
CrossRef - Zhang X., Yu L., Shi J., Li S., Yang S., Gao W., … Geng, C. Antibiotics modulate neoadjuvant therapy efficiency in patients with breast cancer: A pilot analysis. Scientific Reports. 2021;11(1):1-7.
CrossRef - Salek RM, Maguire ML, Bentley E, Rubtsov DV, Hough T, et al. A metabolomic comparison of urinary changes in type 2 diabetes in mouse, rat, and human. Physiol Genomics. 2007; 29: 99–108.
CrossRef - Adams S.H., Hoppel C.L., Lok K.H., Zhao L., Wong S.W., et al. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid beta-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African-American women. J Nutr 2009;139: 1073–1081.
CrossRef
Accepted on: 08 Dec 2023
Second Review by: Amany A Salama
Final Approval by: Dr. Nurul Huda





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








