Characteristics of Inoculated Multi-Stage Apple Fermentation Over Spontaneous Fermentation for the Production of Traditional Functional Beverage Jiaosu: Antioxidant, Antibacterial Activities and Metabolic Changes
Shanshan Han*
, Hong Yin and Qiang Zhang Bio-Agriculture Institute of Shaanxi, Shaanxi Academy of Sciences, Xianning Road Nr 125, Xincheng District, Xi’an, China.
Corresponding Author Email: 492748940@qq.com
DOI : http://dx.doi.org/10.12944/CRNFSJ.9.2.24
Download this article as:
![]()
The characteristics of fermented apple juice ‘Jiaosu’ as a traditional folk beverage were determined. Sequential alcoholic, acetic and malolactic fermentation was mediated either by intrinsic or inoculated microbiota over six months at room temperature under static conditions. The fermentation mixture was composed of fruits of Malus pumila Mill., var. Fuji (Rosaceae) and fruit of Citrus limon Burm. f., var. Eureka (Rutaceae), acacia honey of Robinia pseudoacacia L. (Fabaceae) and spring water at a weight ratio of 3:1:10. Product anti-oxidative features, antimicrobial capacities and the metabolic byproducts were determined at the end of fermentation. Our results showed an elevation of antioxidant activity, good antimicrobial ability and enrichment of bioactive compounds in the directed fermentation inoculated with Saccharomyces cerevisiae, Acetobacter pasteurianus and a mixture of Lactobacillus acidophilus, Lactobacillus plantarum and Lactobacillus paracasei compared to the spontaneous fermentation. Prebiotic fermentation tended to preserve and even improve the antioxidant capacity throughout the fermentation process and generated valuable substances. Though it may reduce the amounts of some other bioactive compounds, the final product quality of directed fermentation was reliable. This study provides valuable information for understanding tradition functional beverages based on fermented fruits.
KEYWORDS:Antioxidant; Apple; Functional Beverage; Jiaosu; Metabolites; Spontaneous Fermentation
Introduction
Fresh fruits and vegetables are rich in antioxidants, vitamins, dietary fibers, minerals, phenolic compounds, and other biochemicals.1 Fermentation transforms those compounds into possible prebiotics and probiotics, as well as enhancing nutrient bio-accessibility and bioavailability, leading to increased health and nutritive value. Moreover, fermented beverages contain less anti-nutritional factors and instead have more enhanced bioactive molecules generated by fermentation.2 Traditional fermented beverages are not only in great demand with consumers who wish to abstain from alcohol or vegetarians, but also are increasingly being recognized by the general public as healthy alternatives. Notably, they have high antioxidant activity and a correspondingly high radical scavenging capacity.3,4
Practices of fruit and vegetable fermentation in Asia started in ancient times. As a traditional folk beverage, ferment fruits and vegetables are not only served as daily food supplements but also considered to contribute to human health in Asian culture. Their use is passed across generations as regional tradition,5 including the known kimchi6 (Korean fermented vegetables), miso7 (Japanese fermented soybeans), kefir8 (fermented milk with Caucasian and Tibet origin), kombucha9 (sweet and sour drink made with fermented tea) and Jiaosu10 (fermented fruit and vegetable juice). Naturally fermented foods and alcoholic beverages rely on autochthonous microbiota, such as lactic acid bacteria (LAB), or Saccharomyces and other yeasts that naturally colonize fruits and vegetables.11,12 Among them, Lactobacilli are the most known probiotics. One-step lactic acid fermentation is a favored method for the manufacture of probiotic-rich products that can improve digestion and nutrient assimilation. More importantly, it can improve product palatability. Among LAB, L. plantarum is of great interest and frequently employed in fruit and vegetable fermentation due to its production of high-added value compounds.1 Individual LAB strains or their combinations can impart different flavors and textural quality to fermented products. Generally, L. plantarum, L. rhamnosus, L. gasseri, and L. acidophilus were reported as good fermentation starters.13
Apple skin and flesh contain large amounts of phenolic compounds. Moreover, there is evidence that the consumption of dry apples, apple skin, apple juice, apple polyphenol extracts, has a hypolipidemic effect.14 Corresponding clinical studies indicate that the fiber fraction, and especially pectin, as well as polyphenols are the main beneficial constituents.14,15 Consequently, apples are considered to be a good resource for LAB growth to make functional juice.16 To commercialize homemade fermented beverages based on apples and autochthonous microbiota, recent efforts have been made to develop industrial processes that utilize a microbial consortium including yeasts, acetic acid bacteria and lactic acid bacteria. There has also been a commercialization effort to label such products as functional beverages due to their putative health benefits. However, the rapid development and lack of proper technical background or scientific knowledge left producers and customers without proper understanding beyond the marketing narrative. In this study we investigated the effects of spontaneous and directed fermentation on metabolic byproducts, anti-oxidative features and antimicrobial activities during the fermentation of traditional apple beverage. This work contributes to the commercial development and utilization of traditional fermented fruit beverages.
Materials and Methods
Micro Organisms and Starter Cultures
S. cerevisiae Hansen137, purchased from Guangdong Microbial Culture Collection Center (Guangdong, China), was cultured at 28 °C and 200 rpm for 48 h. Pure culture of A. pasteurianus BNCC335801 from BeNa Culture Collection (Beijing, China) was grown at 30 °C and 200 rpm for 24 h on a medium consisting of 5 mL ethanol, 10 g yeast extract, and 10 g glucose per liter. Commercial strains of L. acidophilus, L. plantarum and L. paracasei (VEGE 011 LY0 and 100 DCU) from DANISCO were grown in MRS medium to 1´106 CFU/mL to start the lactic acid fermentation. The same procedure of draining growing culture and resuspending cell pellet into 10 mM PBS buffer was used for sequentially inoculating yeast, acetic bacteria and lactic bacteria in directed fermentation. Each fermentation stage was statically incubated at room temperature with 1% inoculation.
Preparation of Traditional Apple Beverage
Here, Fuji apples from Luo Chuan City, Eureka lemon fruits and RP acacia honey (sugary content 89.7 g/100g, producer: Guan Sheng Yuan, Shanghai) were purchased from a local supermarket in Shaanxi, China, rinsed with sterile water and dried thoroughly. We chopped the apples into small pieces and removed the pits. The Eureka lemons were steeped in boiling water, drained, sliced, and the seeds removed. Finally, 1.44 kg of fruit (Fuji apples 80% + Eureka lemons 20%), 0.48 kg of RP honey and 4.8 kg of spring water were mixed in an 8-liter tank at a weight ratio of 3:1:10, yielding a total of 6.72 kg per tank. Eureka lemon slices were placed on the top layer of every single tank to slow down apple discoloration. The tank is a glass container (diameter: 21 cm, height: 30.5 cm) with an airlock system, a temperature meter, and a sampling valve with in-built filter. A total of 12 tanks were set up for two treatments of static spontaneous and static directed fermentation. Two groups were named N for spontaneous fermentation and group D for directed fermentation. Six independent biological replicates were grouped for one treatment. The airlock on the tank was open for the first 30 days of ethanol fermentation and second 30 days of acetic acid fermentation. From day 61 onwards, the airlock was closed until the end of the fermentation (150 d). No pasteurization or filtration was carried out in-between two fermentation stages. The fermentation tank and preparation room were fumigated with ozone to minimize the risk of contamination.
Sample Collection During the Fermentation
The room temperature was maintained at 20 ± 2 ℃. The glass tanks were shaken thoroughly before taking samples periodically during the ethanol fermentation, acetic acid fermentation and lactic acid fermentation. Corresponding sampling times and group names were used as follows: f.x. NovN stands for sample taken in November from group N. Overall, there were samples NovN, DecN, FebN, AprN for spontaneous fermentation, and NovD, DecD, FebD, AprD for directed fermentation. A sample volume of 55 mL was filtered out at each time point. The sample preparation procedure before analysis was done as follows: 2 mL of subsamples were centrifuged at 13,000 × g for 5 min. The supernatants were then stored at −80 °C for further metabolite analysis. 50 mL of subsamples were used for measuring the anti-oxidative activity at the indicated time points. To determine the antimicrobial properties, 3 mL of subsamples were filtered to eliminate endogenous microbes.
In Vitro Anti-Oxidative Assays
The antioxidant activity of ferment apple juice was characterized via a 2,2-diphenyl-1-picrylhydrazyl radical (DPPH) free radical scavenging assay, hydroxyl radical (•OH) assay, superoxide dismutase (SOD) activity measurement and total phenolic content determination.
The DPPH scavenging activities were measured as follows: 2.0 mL of 0.04 mg/L fresh DPPH ethanolic solution (Shanghai Macklin Ltd, China) was added to a mixture of 1 mL sample, 1 mL double distilled water (ddH2O), and incubated for 30 min in dark. The absorbance of the mixture was measured at 517 nm and blanked with the control. The control was carried out with water or ethanol instead of the tested sample, and methanol instead of DPPH was used as blank. The radical scavenging ability was shown as % inhibition per 0.1 mL using the formula: Inhibition%= (control absorbance – sample absorbance)/ control absorbance × 100.
Scavenging potential of •OH was measured using the Fenton reaction as described by Smirnoff and Cumbes (1989). The 3.0 mL reaction mixture contained 1.0 mL of 1.5 mM FeSO4, 0.7 mL of 6 mM hydrogen peroxide, 0.3 mL of 20 mM sodium salicylate (Lingfeng, Shanghai) and 0.3 mL of the sample. After incubation for 1 h at 37 °C, the absorbance of the hydroxylated salicylate complex was measured at 510 nm. The •OH quenching activity (E%) was calculated using the equation: E (%) =[Ax-(Ay-Az)] ÷Ax×100, where Ax is the control absorbance of ddH2O (without sample), Ay is the absorbance in the presence of sample, and Az is the absorbance without sodium salicylate.
SOD activity was measured according to national guidelines in China (GB/T5009.171-2003) using the pyrogallol autoxidation method monitored spectrophotometrically at 325 nm. Inhibition of pyrogallol reduction to 50% of maximal is defined as 1 U of SOD activity, and enzyme activity was expressed in units per mL sample.
The total phenolic (TP) content was measured using the Folin–Ciocalteu method. Gallic acid (Tanmo Quality Inspection Ltd. Beijing, China) was used as the reference compound to calibrate the TP content. The standard curve was constructed using gallic acid in 95% ethanol at various concentrations. The Folin–Ciocalteu reaction included 0.1 mL sample, 0.9 mL ddH2O, and 2 mL Folin–Ciocalteu reagent. After 3 min, 2 mL of 10% (w/v) Na2CO3 was added into the reaction mixture and incubated for 1 h at 25 °C. The absorbance of the mixture was measured at 760 nm against distilled water on a UV spectrophotometer. The TP content was expressed as μg/mL gallic acid equivalents.
In Vitro Antibacterial Potential of Fermented Apple Juice
Bacterial inhibition was determined using a method described in 2013.17 Four food-borne pathogenic microorganisms, Staphylococcus aureus (Sau, ATCC25923), Escherichia coli (Ec, ATCC25922), Salmonella typhimurium (Sty, ATCC14028) and Shigella flexneri (Sfl, ATCC12022), were purchased from Hope Biosciences in Qingdao, China. A stock culture of each strain was prepared according to the guidelines provided by the supplier. The antibacterial effect was quantified by measuring the diameters of the inhibition zones formed around the discs (diameter: 7 mm) using a caliper.
Metabolic Profiling of Apple Fermentation Using Non-Targeted Metabolomics
For each group, intracellular metabolites were extracted from six independent biological replicates and analyzed using gas chromatography / time-of-flight–mass spectrometry (GC/TOF–MS) provided by BMKCloud (www.biocloud.net). An Agilent 7890 gas chromatograph system coupled with a Pegasus HT time-of-flight mass spectrometer was used. The capillary column (30 m length × 0.25 μm internal diameter × 0.25 μm film thickness) was from J&W Scientific, US. The initial oven temperature was 50 °C for 1 min, followed by ramp to 310 °C for 8 min at a rate of 20 °C/min. Inlet flow was 3 mL/min. Temperatures of the injector, transfer line, and ion source were 280, 280, and 250 °C, respectively. The energy was −70 eV in electron impact mode. The scan mode was set to an m/z range of 50 – 500 at a rate of 20 spectra/s. LECO ChromaTOF software (version C; LECO, St. Joseph, MI, USA) was used for detection of peaks and deconvolution of mass spectra. Mass data were further processed using the Fiehn library and the NIST library.
Statistical Analysis
Antioxidative properties and pathogen inhibition were measured in three out of six independent replicates and each measurement was shown as Mean ± Standard Deviation (SD) (N=2). GraphPad Prism 8.0 was used to perform the analysis. The significance of differences was calculated by one-way analysis of ANOVA using Tukey’s post-hoc test at p < 0.05. Metabolites were assessed based on six independent replicates. Principal component analysis (PCA) was used to investigate the relationship between samples after fermentation using R software. An OPLS-DA (orthogonal projections to latent structures discriminant analysis) model was used to further validate the sample separation.
Results and Discussion
The present study investigated three stages of directed fermentation in making traditional functional beverage. Different to the natural fermentation method, the first stage was the growth of S. cerevisiae on sugar and ethanol production for 30 days, followed by growth of A. pasteurianus and production of acetic acid for the next 30 days, converting ethanol to acetic acid in the presence of oxygen. Afterwards, the airlock was closed, allowing chelation and esterification to proceed for 30 days under anaerobic conditions. From day 91, the predominant metabolic activity was the conversion of malic acid into lactic acid during malolactic fermentation, along with the general maturation process. Spontaneous fermentation relies on the indigenous flora present on the surface of the apples. The fermented juice in both groups presented a pleasant aroma and taste. The final pH of all the cultures was 3.41 ± 0.12. The final ethanol content was 0.258 ± 0.16 g/100g for group N and 0.151 ± 0.09 g/100g for group D. The residual sugar was below 2 g/L in all cases. The room temperature was kept between 18-20 °C throughout the fermentation period for better aroma development.18
Antioxidant Activities of Fermented Apple Beverage
Among the methods for testing the antioxidant ability, DPPH assay is now widely used in many matrices such as juice fruit,19,20 vegetable oils,21 and propolis,22 due to its simplicity, rapidity, sensitivity and reproductivity. Considering that polyphenols are the main potential antioxidant substances accounted for a good radical scavenging ability in fresh apples and their relavent products23, we determined the antioxidant capacity of the apple beverage using DPPH and •OH scavenging assay, SOD activity and TP content.
An increase in the DPPH value was observed as the fermentation started. Directed fermentation exhibited a steady increase of DPPH radical potential compared to spontaneous fermentation (Fig.1a). A total rise of 38.863 ± 0.912% was observed in the DPPH radical potential in group D, compared to 32.554 ± 1.365% in the control group (p<0.05). This result indicated that the overall antioxidant capacity of group D was higher than that of group N.
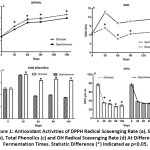 |
Figure 1: Antioxidant Activities of DPPH Radical Scavenging Rate. |
Polyphenol hydrolyzation can lead to an increase of antioxidant activity.24 This was reflected clearly in group N (Fig.1c). The TP reached a maximal value of 370 μg/mL in the directed fermentation and 364.3 μg/mL in the spontaneous fermentation at day 30. Subsequently, the TP value started to decrease, with a slope of -49.33 for group D and -173.67 for group N. The TP level dropped greatly in the naturally fermented group N, with less fluctuation observed in group D (Fig.1c, p<0.05), where a slight drop occurred in TP at day 60 and remained stabled above 300 μg/mL until the end of fermentation. The high phenolic content in group D could be a result of depolymerization of conjugated phenolic compounds by (poly)phenoloxidases,25 exopolysaccharides released by LAB fermentation,26 efficient extraction and transformation of phenolics by microorganisms under the ‘skin-flesh’ maceration procedure,27 or a combination of all of the above. Similar results were obtained in other fermentation studies with milk and juice.28-30
Superoxide radicals and •OH are among the most important reactive oxygen species in biological systems. SOD is the main scavenger of superoxide radicals, which are the precursor of •OH. In this sequential scheme of O2 ® superoxide radicals31® •OH, SOD inhibits the accumulation of superoxide radicals, thus eliminating the generation of •OH. This explains why the increase of SOD was related with a corresponding fall in •OH % levels. The directed fermentation showed lower SOD activity than in control group (Fig.1b, p<0.05). Yeast inoculation quickly started the ethanol fermentation and consumed the oxygen. Consequently, less SOD was needed in group D and more superoxide radicals were generated in group N. Accordingly, slow oxygen consumption with no inoculation (group N) generated more superoxide radicals. The SOD activity decreased after an initial increase in the first 30 days of fermentation in both cases. This reduction may be due to the acidification of the medium due to the acetic and lactic fermentation, the low temperature used for the fermentation, or a stressful growth environment.32 The phenomenon of lower TP and higher SOD activity in the spontaneously fermented group may be a result of LAB growth.16 LAB can degrade phenols, and some compounds in the inoculated group may inactivate SOD activity. In agreement with this hypothesis, a slight increase in SOD activity was observed following the LAB fermentation. This may be the result of lower •OH production and accumulation of SOD from LAB. The inhibition of •OH formation was clearly seen starting from 77.13% to 29.17±1.827% and 23.52±1.716% for group D and N, respectively (Fig.1d, p<0.05). This change occurred mainly during the ethanol fermentation and remained stable until the end of fermentation. A greater reduction of •OH was seen in the directed fermentation group.
In general, a similar development trend for all selected parameters was observed during the entire fermentation period (p<0.05). The change trend of DPPH radical scavenging was reversely associated with reduced •OH levels (r=0.922, p=0.026). TP and •OH % were strongly negatively correlated, with an r value of 0.824. There was a moderate positive correlation between TP and DPPH% (r =0.563). The correlation between TP and SOD had an r value of 0.414. SOD is an intracellular enzyme, and its activity can be influenced by many factors. In this case, SOD showed the lowest correlation with the other three indicators. It has been reported that the total antioxidant activity of a culture is not only related to the TP,4 but also influenced by the fermentation process, which involves microorganisms and their metabolites.33 However, a recent study found that most organic acids and other fermentation metabolites do not contribute to the antioxidant capacity.34 Therefore, the total antioxidant value of ferment juice was sufficiently reflected by the DPPH and TP measurements. The high values of DPPH and TP in group D suggested that sequential inoculation is a good method for preserving and even improving the antioxidant activity of the fermented beverage.
Antibacterial Activity of the Fermented Juice Against Foodborne Pathogens
Traditional fermented apple juice making uses unpasteurized juice. However, certain microorganisms present in unpasteurized juice can cause possible food spoilage and food-borne illnesses. Consideration was thus given to the anti-bacterial activity of the samples after fermentation. Four potential contaminant microbes (Sau, Ec, Sty and Sfl) were tested and the inhibitory effects are shown in Fig.2 (p<0.05). The growth inhibition of Sau, Sty and Sfl was weaker in the directed fermentation group than in the spontaneously fermented control group, while greater inhibition against Ec growth was shown with the inoculated group. Both fermentation treatments (pH 3.41 ± 0.12) increased the inhibitory effects against pathogens compared to unfermented apple juice (pH 5.53). Considering that acidity may affect the inhibition result, the samples were neutralized to pH 7 and growth inhibition was reduced accordingly for both fermentation groups. The inhibition zones were all smaller than 8 mm. Low sample pH and antimicrobial compounds in the two fermentation groups were considered to be the main inhibitory factors. This result is in line with reports that unpasteurized fermented beverages are generally considered as a safe product due to the presence of ethanol, low pH, bacteriocins and other antibacterial/antifungal compounds, as well as organic acids from lactic acid fermentation.35,36
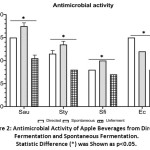 |
Figure 2: Antimicrobial Activity of Apple Beverages from Directed Fermentation and Spontaneous Fermentation. |
It should be noted that we only tested four potentially pathogenic bacteria, and other species deserve attention in future studies, such as acid tolerant enterohemorrhagic E. coli (EHEC) 0157:H7.37 Further microbial safety studies are needed to examine or eliminate foodborne pathogens in traditional fermented apple juice before consumption.
Overview of Metabolic Changes During the Fermentation
PCA and orthogonal projections to latent structures discriminant analysis (OPLS-DA) are normally used to construct a map of the general relationship among different samples. The PCA plot shows the time-dependent change of metabolite profiles during the entire fermentation (Fig.3). Three selected time points (DecD, AprD, AprN) were well discriminated. Metabolite dots from either of the samples did not overlap, indicating an alteration over time in both fermentation settings. Six biological replicates are shown for each time point. Two selected samples from the directed fermentation, at day 30 and day 150, were marked as DecD and AprD. Spontaneous fermentation at 150 d was marked as AprN. A larger deviation can be seen among the six replicates of AprN, as expected. After 150 days of fermentation, AprD and AprN were separated by PC2. The two principal components (PC1 and PC2) explained 95.7% of the total data variance.
The OPLS-DA score plot based on non-volatile metabolite profiles was further constructed and validated by permutation testing. When comparing AprN versus AprD and DecD versus AprD, the Q2 calculated based on the cross-validation was 0.836 and 0.977, respectively, indicating that the supervised models have good predictability. R2X and R2Y are parameters calculated from cumulative modeled variations in the X and Y matrix. The R2X and R2Y of AprN versus AprD were 0.364 and 0.836, compared to 0.576 and 0.999 when valuating DecD versus AprD, indicating a more robust model in the latter. From Fig.3 and the OPLS-DA scores, we could see that the directed fermentation process offered a more predictable and controllable final product, whereas spontaneous fermentation created a larger separation among its six biological replicates.
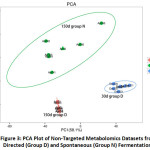 |
Figure 3: PCA Plot of Non-Targeted Metabolomics Datasets from Directed (Group D) and Spontaneous (Group N) Fermentation. |
A total of 40 detected compounds were found when comparing AprN to AprD. After 150 days of fermentation, 21 of these 40 compounds increased and 19 decreased. Fig.4A shows the fold-change analysis of the top 10 compounds with the largest differences between AprN and AprD. Of note was the amount of capric acid and quinic acid in group D after fermentation, with a relative concentration of 0.83 ± 0.16 g/100 g sample and 1.39 ± 0.08 g /100 g sample (Fig.4C). As one of the healthy medium-chain fatty acids, capric acid exhibits antimicrobial and anti-inflammatory activities.37,38 Notably, its content increased the most after fermentation. Quinic acid is reported to have strong antibacterial activity,39 and its content was dramatically increased in the fermentation broth. Additionally, the concentration of lactic acid was 15.8 times greater than in the control.
 |
Figure 4: Metabolic Changes After Fermentation. |
Among the 132 differentially abundant compounds identified between ethanol fermentation (DecD) and three steps fermentation (AprD), 38 compounds were increased and 94 decreased. As shown in Fig.4B, glucuronic acid showed the greatest increase after acetic and lactic fermentation, with a relative concentration of 4.19 ± 0.23 g/100 g sample. Glucuronic acid is produced by the natural symbiosis of bacteria and yeasts in the fermentation process, and it has a positive effect on human health.40,41 The second compound with a large change was hydroxyphenyllactic acid (relative concentration 1.98 ± 0.14 g/100 g sample). Its production can be attributed to the inoculation with Acinetobacter and Lactobacillus.42 Microbially produced hydroxyphenyllactic acid decreases reactive oxygen species.43 The metabolic intermediate 9-fluorenone was accumulated together with lysine and galactinol under this serial fermentation strategy. Their potential favorable antimicrobial effects have already been described elsewhere, as well as their roles as essential amino acids and antioxidants.44,45 In agreement with a previous study,46 the lysine concentration increased to 0.85 ± 0.24 g/100 g sample after 30 days of acetic fermentation following the alcoholic fermentation. Additionally, epicatechin, inositol, oxamic acid, dihydroxybenzoic acid, galacturonic acid decreased to a certain degree. The overall health benefits of these fermentation products need to be further evaluated in future studies.
Conclusions
Consumer interest in functional fermented beverages is growing rapidly. Numerous microbial combinations, with unclear effects on product quality, diverse production procedures and other undefined factors all hinder the development of standardized functional beverages. Consequently, systematic studies are needed to enable informed production and build consumer confidence. With this in mind, we provide a general microbiological and metabolomic outline of a traditional functional beverage based on fermented apple juice. We developed a simple protocol with three fermentation steps. Following a traditional method where maceration of unpasteurized fruit skin and flesh was applied, our fermentation process was extended over months. Intrinsic and inoculated microflora in this case mediated the whole fermentation process. The presence of alcoholic, acetic and malolactic fermentation was beneficial for the overall antioxidative activity and pathogen inhibition. Overall, the inoculated three-stage fermentation led to an enrichment of bioactive metabolites in fermented apple juice over its spontaneously fermented control. Thus, this fermentation approach can be used to develop improved health drinks for the Asian market, as well as an alternative for fresh fruit preservation. Our findings provide a scientific basis for understanding traditional Asian fermented fruit beverage Jiaosu.
Author Contribution
Conceptualization, S.H. and Q.Z.; methodology, S.H. and H.Y.; resources, S.H. and H.Z.; writing—original draft preparation, S.H.; writing—review and editing, Q.Z.; project administration and funding acquisition. All authors have read and agreed to the publication of the final manuscript.
Conflict of Interest
No potential conflict of interest was reported by the authors. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Funding Sources
Shaanxi Academy of Sciences [Grant No. 2018nk-05 and Y18N001G18] supported the present work.
Acknowledgment
We thank for Jianxin Tan from Shaanxi Xinsheng Jixiang technology co., Ltd, for his input to the project.
References
- Septembre-Malaterre A., Remize F., Poucheret P. Fruits and Vegetables, as A Source of Nutritional Compounds and Phytochemicals: Changes in Bioactive Compounds During Lactic Fermentation. Food Res Int. 2018; 104: 86–99.
CrossRef - Gan R. Y., Shah N. P., Wang M. F., Lui W. Y., Corke H. Lactobacillus plantarum WCFS1 Fermentation Differentially Affects Antioxidant Capacity and Polyphenol Content in Mung Bean (Vigna radiata) and Soya Bean (Glycine max) Milks. J Food Process Pres. 2017; 41(1): e12944.
CrossRef - Cardoso A., Lemos L., Frighetto M., Nunes E. Total Phenolics Extracted from the Skin of Fuji Apple and Incorporated by Liposome in Galenic Bases : An Alternative to Use By-products of Food Industry. Rom Biotech Lett. 2011; 16: 40–45.
- Chinnici F., Bendini A., Gaiani A., Riponi C. Radical Scavenging Activities of Peels and Pulps from cv. Golden Delicious Apples as Related to Their Phenolic Composition. J Agric Food Chem. 2004; 52(15): 4684–4689.
CrossRef - Swain M. R., Anandharaj M., Ray R. C., Rani P. R. Fermented Fruits and Vegetables of Asia: A Potential Source of Probiotics. Biotechnol Res Int. 2014; 1–19.
CrossRef - Park K., Jeong J. K., Lee Y. E., Daily J. W. Health Benefits of Kimchi (Korean Fermented Vegetables) as A Probiotic Food. J Med Food. 2014; 17(1): 6-20.
CrossRef - Ito K. Review of the Health Benefits of Habitual Consumption of Miso Soup: Focus on the Effects on Sympathetic Nerve Activity, Blood Pressure, and Heart Rate. Environ Health Prev Med. 2020; 25(1): 45.
CrossRef - Rosa D. D., Dias M. M. S., Grześkowiak Ł.M., Reis S.A., Conceição L.L., Peluzio M. D. C. G. Milk Kefir: Nutritional, Microbiological and Health Benefits. Nutr Res Rev. 2017; 30(1): 82-96.
CrossRef - Villarreal-Soto S. A., Beaufort S., Bouajila J., Souchard J. P., Taillandier P. Understanding Kombucha Tea Fermentation: A Review. J Food Sci. 2018; 83(3): 580-588.
CrossRef - Dai J., Sha R., Wang Z., Cui Y., Fang S., Mao J. Edible Plant Jiaosu: Manufacturing, Bioactive Compounds, Potential Health Benefits, and Safety Aspects. J Sci Food Agric. 2020; 100(15): 5313-5323.
CrossRef - Deshpande M.V. Microbial Diversity on Grapes and Other fruits: Role and Significance in Fermentation. In: Satyanarayana T., Das S., Johri B. (eds) Microbial Diversity in Ecosystem Sustainability and Biotechnological Applications. Singapore: Springer; 2019: 187-204. https://doi.org/10.1007/978-981-13-8487-5_8.
CrossRef - Tamang J. P., Shin D. H., Jung S. J., Chae S. W. Functional Properties of Microorganisms in Fermented Foods. Front Microbiol. 2016; 7: 578.
CrossRef - Karovičová J., Drdák M., Greif G., Hybenová E. The Choice of Strains of Lactobacillus Species for the Lactic Acid Fermentation of Vegetable Juices. Eur Food Res Technol. 1999; 210(1): 53–56.
CrossRef - Koutsos A., Riccadonna S., Ulaszewska M. M., Franceschi P., Trošt K., Galvin A., Braune T., Fava F., Perenzoni D., Mattivi F., Tuohy K. M., Lovegrove J. A. Two Apples A Day Lower Serum Cholesterol and Improve Cardiometabolic Biomarkers in Mildly Hypercholesterolemic Adults: A Randomized, Controlled, Crossover Trial. Am J Clin Nutr. 2020; 111(2): 307–318.
CrossRef - Shahrokh K., Rong T., Djamila R., Raymond Y., Marie C. T., Rupasinghe H. P. V. Polyphenol Composition and Total Antioxidant Capacity of Selected Apple Genotypes for Processing. J Food Compost Anal. 2008; 21(5): 396–401.
CrossRef - Espirito-Santo A. P., Carlin F., Renard C. M. G. C. Apple, Grape or Orange Juice: Which One Offers The Best Substrate for Lactobacilli Growth? – A Screening Study on Bacteria Viability, Superoxide Dismutase Activity, Folates Production and Hedonic Characteristics. Food Res Int. 2015; 78: 352–360.
CrossRef - Wen Y. L., Yan L. P., Chen C. S. Effects of Fermentation Treatment on Antioxidant and Antimicrobial Activities of Four Common Chinese Herbal Medicinal Residues by Aspergillus oryzae. J Food Drug Anal. 2013; 21(2): 219–226.
CrossRef - Peng B., Li F., Cui L., Guo Y. Effects of Fermentation Temperature on Key Aroma Compounds and Sensory Properties of Apple Wine. J Food Sci. 2015; 80(12): S2937–S2943.
CrossRef - Giuffrè A. M. Bergamot (Citrus bergamia, Risso): The Effects of Cultivar and Harvest Date on Functional Properties of Juice and Cloudy Juice. Antioxidants. 2019; 8(7): 221.
CrossRef - Londoño M. Z., Chaparr D.,. Rojan B. A., Arbelae A. F. A., Betancu L. F. R., Celis M. E. M. Effect of Storage Time on Physicochemical, Sensorial, and Antioxidant Characteristics, and Composition of Mango (cv. Azúcar) Juice. Emir J Food Agric. 2017; 29(5): 367-377.
- Giuffrè A. M., Caracciolo M., Zappia C., Capocasale M., Poiana M. Effect of Heating on Chemical Parameters of Extra Virgin Olive Oil, Pomace Olive Oil, Soybean Oil and Palm Oil. Ital J Food Sci. 2018; 30(4): 715-739.
- Hasan A. E. Z., Mangunwidjaja D, Sunarti T. C., Suparno O, Setiyono A. Investigating the Antioxidant and Anticytotoxic Activities of Propolis Collected from Five Regions of Indonesia and Their Abilities to Induce Apoptosis. Emir J Food Agric. 2014; 26(5): 390-398.
CrossRef - Tarko T., Kostrz M., Duda-Chodak A., Semik-Szczurak D., Sroka P., Senczyszyn T. The effect of apple cultivars and yeast strains on the selected quality parameters and antioxidant activity of fermented apple beverages. CyTA – J Food. 2009; 16(1): 892-900.
CrossRef - Bouayed J., Hoffmann L., Bohn T. Total Phenolics, Flavonoids, Anthocyanins and Antioxidant Activity Following Simulated Gastro-Intestinal Digestion and Dialysis of Apple Varieties: Bioaccessibility and Potential Uptake. Food Chem. 2011; 128(1): 14–21.
CrossRef - Svensson L., Sekwati-Monang B., Lutz D. L., Schieber R., Gänzle M. G. Phenolic Acids and Flavonoids in Nonfermented and Fermented Fed Sorghum (Sorghum bicolor (L.) Moench). J Agric Food Chem. 2010; 58(16): 9214-9220.
CrossRef - Yamamoto N., Shoji M., Hoshigami H., Watanabe K., Watanabe K., Takatsuzu T., Yasuda S., Igoshi K., Kinoshita H. Antioxidant Capacity of Soymilk Yogurt and Exopolysaccharides Produced by Lactic Acid Bacteria. Biosci Microbiota Food Health. 2019; 38(3): 97–104.
CrossRef - Way L. M., Jones J. E., Swarts N. D., Dambergs R. G. Phenolic Content of Apple Juice for Cider Making as Influenced by Common Pre-fermentation Processes Using Two Analytical Methods. Beverages. 2019; 5: 1–12.
CrossRef - Chan C. L., Gan R. Y., Shah N. P., Corke H. Enhancing Antioxidant Capacity of Lactobacillus acidophilus-Fermented Milk Fortified with Pomegranate Peel Extracts. Food Biosci. 2018; 26: 185–192.
CrossRef - Kwaw E., Ma Y., Tchabo W., Apaliya M. T., Wu M., Sackey A. S., Xiao L., Tahir H. E. Effect of Lactobacillus Strains on Phenolic Profile, Color Attributes and Antioxidant Activities of Lactic-Acid-Fermented Mulberry Juice. Food Chem. 2018; 250: 148–154.
CrossRef - Vivek K., Mishra S., Pradhan R. C., Jayabalan R. Effect of Probiotification with Lactobacillus plantarum MCC 2974 on Quality of Sohiong Juice. LWT-Food Sci Technol. 2019; 108: 55–60.
CrossRef - Schopfer P., Plachy C., Frahry G. Release of Reactive Oxygen Intermediates (Superoxide Radicals, Hydrogen Peroxide, and Hydroxyl Radicals) and Peroxidase in Germinating Radish Seeds Controlled by Light, Gibberellin, and Abscisic Acid. Plant Physiol. 2001; 125(4): 1591–1602.
CrossRef - Yang X., Zhou J., Fan L., Qin Z., Chen Q., Zhao L. Antioxidant Properties of A Vegetable–Fruit Beverage Fermented with Two Lactobacillus plantarum Food Sci Biotechnol. 2018; 27(6): 1719–1726.
CrossRef - Tarko T., Kostrz M., Duda-Chodak A., Semik-Szczurak D., Sroka P., Senczyszyn T. The Effect of Apple Cultivars and Yeast Strains on Selected Quality Parameters and Antioxidant Activity of Fermented Apple Beverages. CyTA – J Food. 2018, 16(1): 892–900.
CrossRef - Barberis A., Deiana M., Spissu Y., Azara E., Fadda A., Serra P. A., D’Hallewin G., Pisano M., Serreli G., Orrù G., Scano A., Steri D., Sanjust E. Antioxidant, Antimicrobial, and Other Biological Properties of Pompia Juice. Molecules. 2020; 25(14): 3186.
CrossRef - Rather I. A., Seo B. J., Rejish Kumar V. J., Choi U. H., Choi K. H., Lim J. H., Park Y. H. Isolation and Characterization of A Proteinaceous Antifungal Compound from Lactobacillus plantarum YML007 and Its Application as A Food Preservative. Lett Appl Microbiol. 2013; 57(1): 69–76.
CrossRef - Zoghi A., Khosravi-Darani K., Sohrabvandi S., Attar H., Alavi S. A. Effect of Probiotics on Patulin Removal from Synbiotic Apple Juice. J Sci Food Agric. 2017; 97(8): 2601–2609.
CrossRef - Kim H. J., Yoon H. J., Kim S. Y., Yoon Y. R. A Medium-Chain Fatty Acid, Capric Acid, Inhibits RANKL-Induced Osteoclast Differentiation via the Suppression of NF-кB Signaling and Blocks Cytoskeletal Organization and Survival in Mature Osteoclasts. Mol Cells. 2014; 37(8): 598–604.
CrossRef - Huang W. C., Tsai T. H., Chuang L. T, Li Y. Y., Zouboulis C. C., Tsai P. J. Anti-bacterial and Anti-inflammatory Properties of Capric Acid against Propionibacterium acnes: A Comparative Study with Lauric Acid. J Dermatol Sci. 2014; 73(3): 232–240.
CrossRef - Bai J. R., Wu Y. P., Elena G., Zhong K., Gao H. Insight into the Effect of Quinic Qcid on Biofilm Formed by Staphylococcus aureus. RSC Adv. 2019; 9(7): 3938–3945.
CrossRef - Sinir G. Ö., Tamer C. E., Suna S. Kombucha Tea: A Promising Fermented Functional Beverage. In: Grumezescu A. M., Holban A. M. (eds) Fermented Beverages. Volume 5 The Science of Beverages. Woodhead Publishing; 2019: 401–432. https://doi.org/10.1016/B978-0-12-815271-3.00010-5.
CrossRef - Vina I., Linde R., Patetko A., Semjonovs P. Glucuronic Acid from Fermented Beverages: Biochemical Functions in Humans and Its Role in Health Protection. Int J Res Rev Appl Sci. 2013; 14: 217–230.
- Beloborodov N. V., Khodakova A. S., Bairamov I. T., Olenin A. Y. Microbial Origin of Phenylcarboxylic Acids in the Human Body. Biochemistry (Moscow). 2009; 74(12): 1350–1355.
CrossRef - Fedotcheva T. A., Kruglov A. G., Teplova V. V., Fedotcheva N. I., Rzheznikov V. M., Shimanovskii, N. L. Effect of Steroid Hormones on Production of Reactive Oxygen Species in Mitochondria. Biophysics. 2012; 57(6): 792–795.
CrossRef - Choi S. R., Larson M. A., Hinrichs S. H., Narayanasamy P. Development of Potential Broad Spectrum Antimicrobials Using C2-symmetric 9-fluorenone Alkyl Amine. Bioorg Med Chem Lett. 2016; 26(8): 1997–1999.
CrossRef - Nishizawa A., Yabuta Y., Shigeoka S. Galactinol and Raffinose Constitute A Novel Function to Protect Plants from Oxidative Damage. Plant Physiol. 2008; 147(3): 1251–1263.
CrossRef - Roda A., Lucini L., Torchio F., Dordoni R., De Faveri D. M., Lambri M. Metabolite Profiling and Volatiles of Pineapple Wine and Vinegar Obtained from Pineapple Waste. Food Chem. 2017; 229: 734–742.
CrossRef
Accepted on: 04 June 2021
Second Review by: Edhi Nurhartadi
Final Approval by: Prof. Daniel Cozzolino





Web of Science Coverage
Emerging Sources Citation Index (ESCI)
2024 Journal Impact Factor: 1.1
Scopus Journal Metrics
CiteScore 2025: 2.6
CiteScore Details
Sustainable Nutrition: Food Systems, Nutrient Retention, and Public Health Impact
![]()
This journal is a member of, and subscribes to the principles of, the Committee on Publication Ethics (COPE)








